细胞的结构和功能
细胞学说
细胞学说的概述
细胞学说的建立者主要是两位德国科学家施莱登和施旺。后人根据他们发表的研究结果进行整理并加以修正,综合为以下要点:
细胞是一个有机体,一切动植物都由细胞发育而来,并由细胞和细胞产物所构成;
细胞是一个相对独立的单位,既有它自己的生命,又对与其他细胞共同组成的整体生命起作用;
新细胞是由老细胞分裂产生的。
【沪科技版】20 世纪初,美国生物学家威尔逊(E. B. Wilson)曾预测:“一切生物学问题的答案最终都要到细胞中去寻找。”今天,生物学研究的众多重大成果证明了这一预测的正确性。
绿眼虫、草履虫等单细胞生物,整个生物体由一个细胞构成。细胞直接与外界非生物环境进行物质和信息交换,生理活动由细胞的不同结构分工完成。例如,草履虫细胞质膜表面凸起形成的纤毛具有运动功能。高等植物由不同器官组成,如根、茎、叶等。人和动物的消化、呼吸、循环等系统也是由相应的器官组成。显微镜下可观察到这些器官都是由无数细胞构成的,形态相似的细胞排列在一起,执行特定的功能。例如小肠绒毛中,柱状的上皮细胞紧密排列在小肠的表面,构成小肠的内表皮,既保护小肠,又可以从肠腔中吸收营养物质;夹杂在上皮细胞中呈酒杯状的是腺细胞,分泌的黏液可以保持小肠的润滑。又如植物叶片中,上表面的栅栏组织细胞排列整齐,充分吸收阳光;背面海绵组织细胞排列疏松,形成空隙,通过气孔与外界相通。由此可见,细胞是生物体结构的基本单位。
细胞学说的建立
细胞学说的建立经历了漫长的过程:
显微镜的发明使人类打开了微观世界的大门,借助显微镜,人们看到了动物体和植物体中各种各样的细胞。
维萨里通过大量的尸体解剖研究,揭示了人体在器官水平的结构。
比夏经过对器官的解剖观察,指出器官由低一层次的结构——组织构成。
罗伯特·胡克用显微镜观察植物的木栓组织,发现这些木栓组织由许多规则的小室组成,他把观察到的图像画了下来,并把“小室”称为细胞(他观察到的只是死细胞)。
列文虎克用自制的显微镜,观察到不同形态的细菌、红细胞和精子等。
马尔比基用显微镜广泛观察了动植物的微细结构,如细胞壁和细胞质。
植物学家施莱登通过对花粉、胚珠和柱头组织的观察,发现这些组织都是由细胞构成的,而且细胞中都有细胞核。在此基础上,他进行了理论概括,提出了植物细胞学说,即植物体都是由细胞构成的,细胞是植物体的基本单位,新细胞从老细胞中产生。
施莱登把他的研究成果告诉了动物学家施旺,施旺很感兴趣并大受启发,决意要证明植物界和动物界这“两大有机界最本质的联系”。施旺主要研究了动物细胞的形成机理和个体发育过程,他认为:动物体也是由细胞构成的,一切动物的个体发育过程,都是从受精卵这个单细胞开始的。
施旺发表了研究报告《关于动植物的结构及生长一致性的显微研究》。施莱登认为新细胞是从老细胞的细胞核中长出来的,或者是在老细胞的细胞质中像结晶那样产生的。
施莱登的朋友耐格里用显微镜观察了多种植物分生区新细胞的形成,发现新细胞的产生原来是细胞分裂的结果。
魏尔肖总结出“细胞通过分裂产生新细胞”、“所有的细胞都来源于先前存在的细胞”。
【浙科版】自然界的生物千差万别、种类繁多,组成这些生物体的细胞形态结构和功能各异,但又具有许多共同的特征。直到 17 世纪中叶,第一台光学显微镜(light mi- croscope)制成后,科学家才发现细胞。因为人眼的分辨率只有约 0.2 mm,而光学显微镜的分辨率可达人眼的 1000 倍,所以利用光学显微镜可以观察到微小的细胞。
1665 年,英国物理学家罗伯特·胡克将软木薄片放在自制的显微镜下观察,发现软木是由许多类似蜂巢的小室构成的。胡克觉得这一形状类似于单人房间,所以他用意为单人房间的 cell 一词命名这些小室为 cella,即细胞。cella 为拉丁语,在英语中仍采用 cell。胡克也成为第一个描述细胞的人。实际上,胡克看到的是已死亡细胞的纤维质细胞壁。
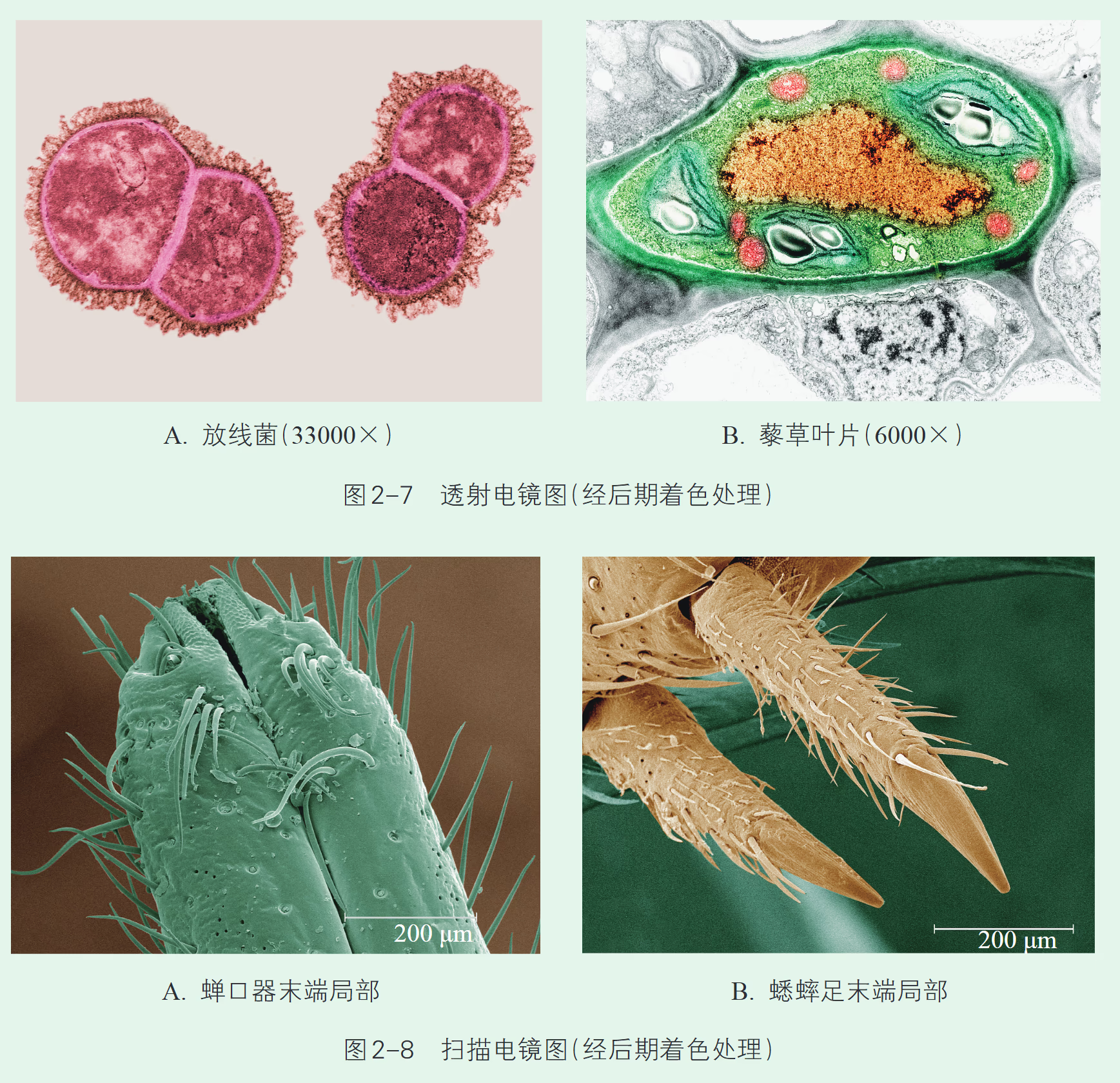
【苏教版】现在已经知道,可见光的波长范围一般为 380~760 nm,因此限制了以可见光为光源的光学显微镜的分辨率,使其最大放大倍数受到很大限制。要提高显微镜的放大倍数,科学家们只能另辟蹊径。20 世纪 30 年代初,德国青年科学家卢斯卡(E. Ruska,1906—1988)等利用高能电子束波长很短的特点,制造出能够放大 10 000 倍的电子显微镜。由于此项贡献,卢斯卡等人获得 1986 年诺贝尔物理学奖。目前使用的电子显微镜已经能把观察的物体放大上百万倍。常用的电子显微镜有透射电子显微镜(transmission electron microscope)和扫描电子显微镜(scanning electron microscope)。
细胞学说的意义
细胞学说揭示了动物和植物的统一性,从而阐明了生物界的统一性(注意不是多样性)。细胞学说中关于细胞是生命活动基本单位的观点,使人们认识到生物的生长、生殖、发育及各种生理现象的奥秘都需要到细胞中去寻找,生物学的研究随之由器官、组织水平进入细胞水平,并为后来进入分子水平打下基础。
【浙科版】细胞学说的提出对近代生物学的发展具有极其重要的意义。它揭示了生物体结构的统一性,揭示了生物间存在一定的亲缘关系,将动物和植物统一起来,为达尔文的进化论奠定了基础。同时,它将人们对生物的研究从宏观领域带入微观领域。对细胞结构的了解,极大地促进了生物学和医学的发展,推进了人类对自然界的认识,有力地促进了自然科学的进步。所以,恩格斯把细胞学说、能量转化与守恒定律和达尔文进化论并列为“19 世纪自然科学的三大发现”。
细胞学说是现代生物学的基础,研究细胞的学科称为细胞学(cytology),是生物学的重要分支。20 世纪 50 年代以后,由于电子显微镜(electron microscope)的使用和相应技术的发展,陆续发现了细胞内许多细微的结构。近年来,由于生物化学和分子生物学的快速发展,细胞学已发展为细胞生物学(cell biology)。
细胞学说是伴随着显微镜技术的发展而不断完善的。在 100 多年的时间里,人们借助显微镜观察了许多动、植物和单细胞的原生动物。人们注意到,自然界的细胞种类极其繁多,形态、体积、功能和生活环境的差异也是巨大的。人类卵细胞直径可达 0.2 mm,可用肉眼直接看到,而某些原始的细胞直径是 0.1 μm,只有通过电子显微镜才能观察到;棉花的一条纤维是单个细胞,长达 3~4 cm;神经细胞的直径不足 1 mm,但长度可超过 1 m;人体细胞的最适温度是 37 ℃,偏离这一温度便会引起身体不适甚至危及生命,而海底热泉附近的某些生物细胞能在 110 ℃以上的高温中生活。即使在同一个个体内,细胞的种类也是多种多样的,如人体中有 200 多种不同类型的细胞,它们分工合作,执行着各自特定的功能。
除病毒外,自然界中的生命都是由细胞构成的。我们可根据细胞是否具有成形的细胞核,将细胞初步分为原核细胞(prokaryotic cell)和真核细胞(eukaryotic cell)。由原核细胞构成的生物称为原核生物(prokaryote),由真核细胞构成的生物称为真核生物(eukaryote)。几乎所有的原核生物都是由单个细胞构成的,如细菌、蓝细菌等。真核生物可分为多细胞真核生物和单细胞真核生物。大多数的动物和植物是典型的多细胞真核生物,草履虫、变形虫、小球藻等是单细胞真核生物。尽管细胞种类繁多,形态各异,但所有细胞都具有相似的基本结构,如细胞膜、细胞质,真核细胞还具有成形的细胞核;所有细胞都具有 C、H、O、N 等基本元素,这些元素组成的无机物和有机物,构成细胞结构,参与细胞生命活动;所有细胞都以 DNA 作为遗传物质。
细胞是一切生命活动的基本单位,即使对于病毒这样的非细胞生物,也只有寄生在活细胞内,才能进行生命活动。对于单细胞生物而言,一个细胞就是一个完整的生命体,生命活动所需的所有代谢活动如消化、排泄和遗传等,均在单个细胞内进行。在多细胞生物体内,功能相同的细胞聚集在一起,共同执行某一功能,如细长的神经细胞集合成束,传导神经信号;扁平的口腔上皮细胞紧密排列,起到保护作用。同时,每个细胞都具有相对独立的代谢系统,机体各项生理功能的完善离不开单个细胞正常的生命活动。
细胞都有相似的基本结构,如细胞膜、细胞质和细胞核,这反映了细胞的统一性。有一类细胞没有成形的细胞核,如大肠杆菌和其他细菌细胞。科学家根据细胞内有无以核膜为界限的细胞核,把细胞分为真核细胞和原核细胞两大类。由真核细胞构成的生物叫作真核生物,如植物、动物、真菌等。由原核细胞构成的生物叫作原核生物。
原核细胞和真核细胞具有相似的细胞膜和细胞质,它们都以 DNA 作为遗传物质,这让我们再一次看到了原核细胞和真核细胞的统一性。
细胞学说中细胞分裂产生新细胞的结论,不仅解释了个体发育,也为后来生物进化论的确立埋下了伏笔。新细胞由老细胞产生,老细胞由更老的细胞产生,如此上溯,现代生物的细胞都是远古生物细胞的后代,小小的细胞内部,凝聚着数十亿年基因的继承和改变。
归纳法是指由一系列具体事实推出一般结论的思维方法。例如,从观察到植物的花粉、胚珠、柱头等的细胞都有细胞核,得出植物细胞都有细胞核这一结论,运用的就是归纳法。归纳法分为完全归纳法和不完全归纳法。根据部分植物细胞都有细胞核而得出植物细胞都有细胞核这一结论,实际上就是运用了不完全归纳法。如果观察了所有类型的植物细胞,并发现它们都有细胞核,才得出植物细胞都有细胞核的结论,就是完全归纳法。科学研究中经常运用不完全归纳法。由不完全归纳得出的结论很可能是可信的,因此可以用来预测和判断,不过,也需要注意存在例外的可能。
个细胞都相对独立地生活着,但同时又从属于有机体的整体功能。单细胞生物能够独立完成生命活动,多细胞生物依赖各种分化的细胞密切合作,共同完成一系列复杂的生命活动。
事实上,动植物以细胞代谢为基础的各种生理活动,以细胞增殖、分化为基础的生长发育,以细胞内基因的传递和变化为基础的遗传与变异,等等,都说明细胞是生命活动的基本单位,生命活动离不开细胞。
无论从结构上还是功能上看,细胞这个生命系统都属于最基本的层次。各层次生命系统的形成、维持和运转都是以细胞为基础的,就连生态系统的能量流动和物质循环也不例外。因此,可以说细胞是基本的生命系统。
【沪科教版】细胞的形态和功能是多种多样的:由于生物的进化、遗传变异和细胞分化,导致构成各种生物体的细胞种类繁多,形态和功能复杂多样。单细胞生物一般是单个细胞独立生活,它们往往有自己特有的形状,如酵母菌大多数呈卵形、圆形或椭圆形。在多细胞生物中,常见的细胞形状有圆形、椭圆形、方形、扁形、梭形、多角形等,也有的形状很不规则。
绝大多数细菌细胞的直径为 0.210μm,而大多数动植物细胞的直径则为 2030μm。不同生物的同种器官或组织,细胞大小总在一个恒定范围之内,不会因生物大小而改变。如人、牛、马、鼠、象的肝细胞大小基本相同。生物体的大小主要取决于细胞的数量,而不是细胞的大小。
细胞形态与功能的统一是细胞的一个重要特征,分化程度较高的细胞尤为明显。例如,动物体内具有收缩功能的肌肉细胞呈长条形或长梭形;人体内有一种神经元,胞体本身的直径一般不超过 10μm,但它伸出去的突起却可达 1m 以上,大大提高了神经冲动传递的效率。高等植物中,对茎部起支持作用的木纤维细胞呈梭形;叶表皮的保卫细胞呈半月形,两个细胞围成一个气孔,有利于控制气体的进出。
原核生物和内共生学说
【浙科版】与真核细胞相比,原核细胞个体微小、结构简单。原核生物由单个原核细胞构成。细菌和蓝细菌属于原核生物。“当我还是个本科生的时候,有两个理论经常被拿出来做反例,以显示科学上的假说可以牵强到何种地步——一个是大陆漂移学说,还有一个就是内共生学说。”一位植物学家回忆说。然而在今天,这两个学说早已被科学界所接受,内共生学说更被看作是对达尔文进化论的重要补充。
美国科学家琳恩·马古利斯(Lynn Margulis,1938—2011,图 2-36)倡导的内共生学说,是关于早期生命演化中细胞如何从简单到复杂的学说。马古利斯认为,真核细胞中线粒体、叶绿体等结构起源于早期的原核生物,某些细菌被原始的真核生物吞噬后,经过长期共生成为线粒体;而蓝细菌被吞噬后逐渐演化为叶绿体。越来越多的证据表明内共生学说是正确的,例如,线粒体和叶绿体内的结构更接近原核生物而不是真核生物等。共生现象在自然界是普遍存在的,如双小核草履虫(Paramecium aurelia)内的某种结构实际上是一种细菌,一些海洋动物如造礁珊瑚与能进行光合作用的虫黄藻(Symbi⁃ odinium)共生。
在地球的生命进化历程中,原核细胞远比真核细胞出现得早。35 亿年前的地层中,就发现了早期蓝细菌的化石。通过对地质岩石分析,可靠的、大量的真核生物化石于 10 亿~8 亿年前才出现。因此,在整个生命进化史前 3/4 的时间里,原核生物是地球上唯一或主要的成员。早期地球没有臭氧层,紫外线很强,大气中 CO2 含量很高,导致地表温度比现在高得多。诸多因素导致早期地球环境并不适合生命生存。地球大气环境的改变,离不开一种结构简单的原核生物——蓝细菌,旧称蓝藻。蓝细菌出现在 35 亿多年前,是最早出现的能进行光合作用的生物。蓝细菌在其出现后的 10 多亿年时间里,一直是地球的主要生物类群。在如此长的时间里,蓝细菌光合作用释放的氧气逐渐在大气圈积累,为真核生物的起源创造了条件,同时促进了臭氧层的形成,阻挡了大量对生物有害的紫外线;蓝细菌的光合作用促使大气圈中 CO2 含量下降,将碳元素转移至岩石圈中形成碳酸盐;CO2 含量的下降逐渐降低了地球表面的平均温度,为陆生生物的出现创造了条件。随着地球环境的改变,逐渐出现原始单细胞真核生物。随着真核生物的繁盛、演化,蓝细菌逐渐衰落,真核生物逐渐占据了生态系统。
没有核膜包被的细胞核,是原核细胞区别于真核细胞最显著的特征。原核细胞的 DNA 较小,位于拟核(nucleoid)或称拟核区,且不以染色质或染色体的形式存在。
原核细胞结构简单,由外向内分别由细胞壁、细胞膜和细胞质构成。原核细胞的细胞壁成分与植物细胞壁不同,对细胞有保护和维持形态的作用。我们生活中服用的不少抗生素,就是通过破坏细菌细胞壁的合成而达到抗菌的作用。细胞壁的外面有时还有一层荚膜,主要由多糖组成,具有保护、黏附等多种功能。有些细菌还有鞭毛,使其能够运动。原核细胞只有核糖体这一类细胞器。
原核细胞的结构虽然简单,但可以完成复杂的生命活动。例如,某些原核细胞可以进行需氧呼吸,与需氧呼吸相关的酶分布于细胞膜和细胞质基质;蓝细菌等原核生物的细胞质含有附着光合色素的光合膜,可以进行光合作用。
原核生物是地球上最早出现的生命形式,其历史可追溯至约 35 亿年前。它们结构简单,分布极广,是生命演化史中的重要基础。原核生物是由原核细胞构成的生物,其最显著的特征是没有以核膜为界限的细胞核,也没有核仁和染色体。现代分类学通常将生物分为真细菌域(Bacteria)、古菌域(Archaea)和真核域(Eukarya)。
- 真细菌:包括常见的细菌、蓝细菌(旧称蓝藻)、放线菌、支原体、衣原体和立克次氏体等。
- 古菌(古细菌):是一类特殊的原核生物,常生活在极端环境(如高温热泉、高盐湖泊)中。其分子生物学特征(如 DNA 复制及转录相关蛋白)有时更接近真核生物。
- 支原体:目前发现的最小、最简单的单细胞生物,其显著特点是没有细胞壁。
结构特征:
拟核(Nucleoid):遗传物质集中分布的区域,没有核膜包被。它通常是一条环状的、裸露的 DNA 分子,不与蛋白质结合形成染色体。
细胞壁:绝大多数原核生物(除支原体外)都有细胞壁,主要成分是肽聚糖。
细胞器:原核细胞非常简单,唯一的细胞器是核糖体,缺乏内质网、高尔基体、线粒体等膜性细胞器。
其他结构:某些细菌具有荚膜(保护作用)、鞭毛(运动器官)或菌毛(附着作用)。
代谢与繁殖:
营养方式多样:
- 自养生物:如蓝细菌(含叶绿素和藻蓝素,能进行光合作用,释放氧气)和硝化细菌(利用化能合成作用制造有机物)。
- 异养生物:绝大多数细菌营腐生或寄生生活。
呼吸类型:包含需氧型(如蓝细菌、硝化细菌,虽然无线粒体,但质膜上附着有呼吸酶)和厌氧型。
繁殖:主要通过二分裂(Binary fission)进行无性繁殖,速度极快。
内共生学说是关于真核细胞起源的最具代表性的假说之一,由美国科学家琳恩·马古利斯(Lynn Margulis)于 1970 年系统论述。该学说认为,真核细胞内的复杂细胞器(主要是线粒体和叶绿体)起源于被原始真核细胞(或古核生物)吞噬的原核生物。
- 线粒体的起源:一种巨大的、具有吞噬能力的原始厌氧细胞吞噬了自由生活的需氧细菌(如变形细菌)。需氧菌未被消化,而是与宿主形成共生关系:需氧菌利用宿主的营养产生能量(ATP),而宿主则在氧气存在的环境下获得了生存优势。最终,这些细菌演化成了线粒体。
- 叶绿体的起源:某些已具有线粒体的早期真核细胞进一步吞噬了自由生活的光合细菌(如蓝细菌)。通过类似的共生进化过程,蓝细菌演化成了专门进行光合作用的叶绿体。
支持内共生学说的主要证据,这些特征使线粒体和叶绿体看起来更像“寄居”在细胞内的细菌:
- 遗传物质的独立性:线粒体和叶绿体拥有自己的环状 DNA 分子,其形状和结构与细菌 DNA 高度相似,且不与组蛋白结合。
- 独立的蛋白质合成系统:它们拥有自己的核糖体(70S),其大小、结构及对某些抗生素(如氯霉素)的敏感性均与细菌一致,而与细胞质中的 80S 核糖体不同。
- 双层膜结构:线粒体和叶绿体具有双层膜。内膜的化学组成与细菌质膜相似(含有褶皱),而外膜则与宿主细胞的质膜(或食物泡膜)更为接近。
- 分裂方式:它们不随细胞核的分裂而分裂,而是像细菌一样通过二分裂进行增殖。
- 共生现象的普遍性:自然界中存在许多现成的实例,如双小核草履虫体内的某些共生细菌,或造礁珊瑚与虫黄藻的共生,证明了这种细胞间的深度协作是完全可能的。
【沪科教版】1905 年,梅列施克夫斯基正式提出真核生物细胞器的共生起源观点,认为叶绿体起源于细胞内共生的蓝细菌。1922 年沃林提出线粒体来自细胞内共生的需氧菌的假说。1970 年,马古利斯出版了《真核细胞的起源》一书,全面系统地论述了内共生假说。她设想,一种厌氧异养原核生物吞入了自由生活的需氧菌,需氧菌没有被消化,两者形成共生关系,需氧菌逐渐进化为真核细胞的线粒体。叶绿体也通过相同的机制由共生的蓝细菌进化而来。通过近几十年的研究,人们找到了越来越多的证据支持内共生假说,主要有:
(1)在形态大小和结构方面,线粒体和细菌相似,叶绿体则与蓝细菌相似。线粒体内膜与细菌质膜都有内褶形成的结构。在化学组成上,线粒体外膜与真核细胞的质膜相似,而线粒体内膜则与原核细胞的质膜相似。
(2)线粒体和叶绿体中都含有 DNA、RNA 和核糖体,是半自主性的细胞器。它们自主生长,都具有合成自身某些蛋白质的能力。
(3)线粒体、叶绿体和原核细胞中所含的 DNA 均为环状分子,不含组蛋白,而真核细胞的核 DNA 则为线状,并与组蛋白结合形成染色体。
(4)细菌、线粒体和叶绿体的蛋白质合成均可受氯霉素和红霉素的抑制,而真核细胞的蛋白质合成对这些抗生素不敏感。反之,亚胺环己酮能抑制真核细胞的蛋白质合成,而对线粒体、叶绿体和细菌的蛋白质合成无抑制效应。
(5)线粒体和叶绿体能以分裂的方式进行增殖,这与细菌的繁殖方式相似,但是它们的分裂与所在细胞的分裂不同步。另外两者均能在异源细胞内长期存在,例如,鸡胚细胞的线粒体引入小鼠体外培养的成纤维细胞后,第四代小鼠细胞内仍可见鸡的线粒体;叶绿体在特殊软体动物细胞内能长期存在。
(6)叶绿体的基因组和光合系统与蓝细菌等原核生物极为相似。
综上所述,线粒体和叶绿体在细胞内表现出较多的自主性与独立性,很多基本特点与细菌较为相似,这似乎指向它们在起源上的联系。但是反对者认为内共生无法解释细胞核的进化过程,也无法解释叶绿体和线粒体基因结构和原核细胞的差异。因此,关于真核细胞的起源,有待今后进一步的研究。
进化的重大意义:
能量革命:线粒体的出现让细胞能进行高效的需氧代谢,为真核生物向多细胞、大型化发展提供了能量保障。
氧气环境的适应:蓝细菌在 35 亿年前开始释放氧气,彻底改变了原始大气。内共生使得真核生物能利用这一环境变化,促进了生物多样性的爆发。
总结来说,原核生物是生命进化的源头,而通过内共生这一跨界的“整合”,简单细胞演变成了高度复杂、分工明确的真核细胞,为后来丰富多彩的生命大爆发奠定了结构基础。
细胞的基本结构
细胞膜的基本认识
细胞膜(cell membrane)也称质膜(plasma membrane),是围绕在细胞外层,将细胞与周围环境区分开的结构。细胞膜是细胞对外界环境的屏障,控制着物质的进出,保证了细胞内部环境的相对稳定,使细胞内的生命活动有序进行。细胞膜(质膜)不仅是细胞的物理边界,更是维持生命活动的关键结构。

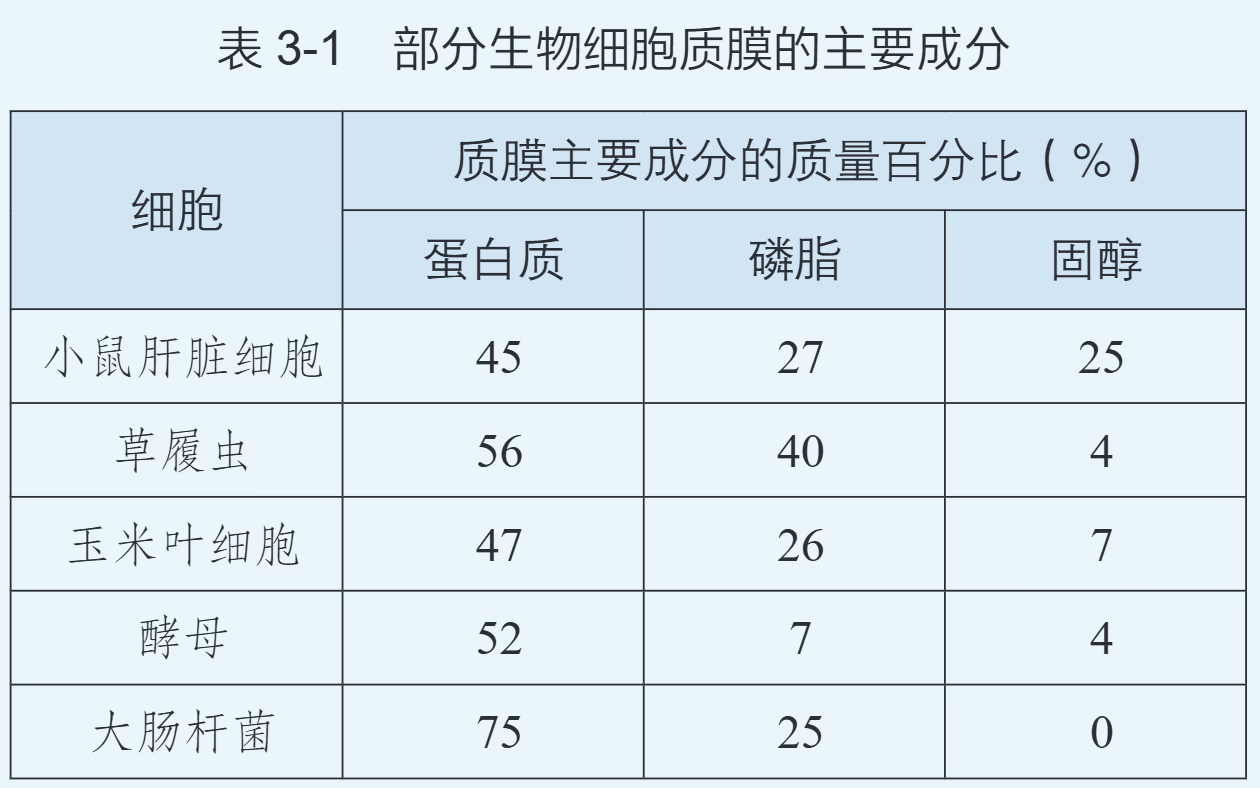
细胞膜主要由脂质和蛋白质组成,含有少量糖类。
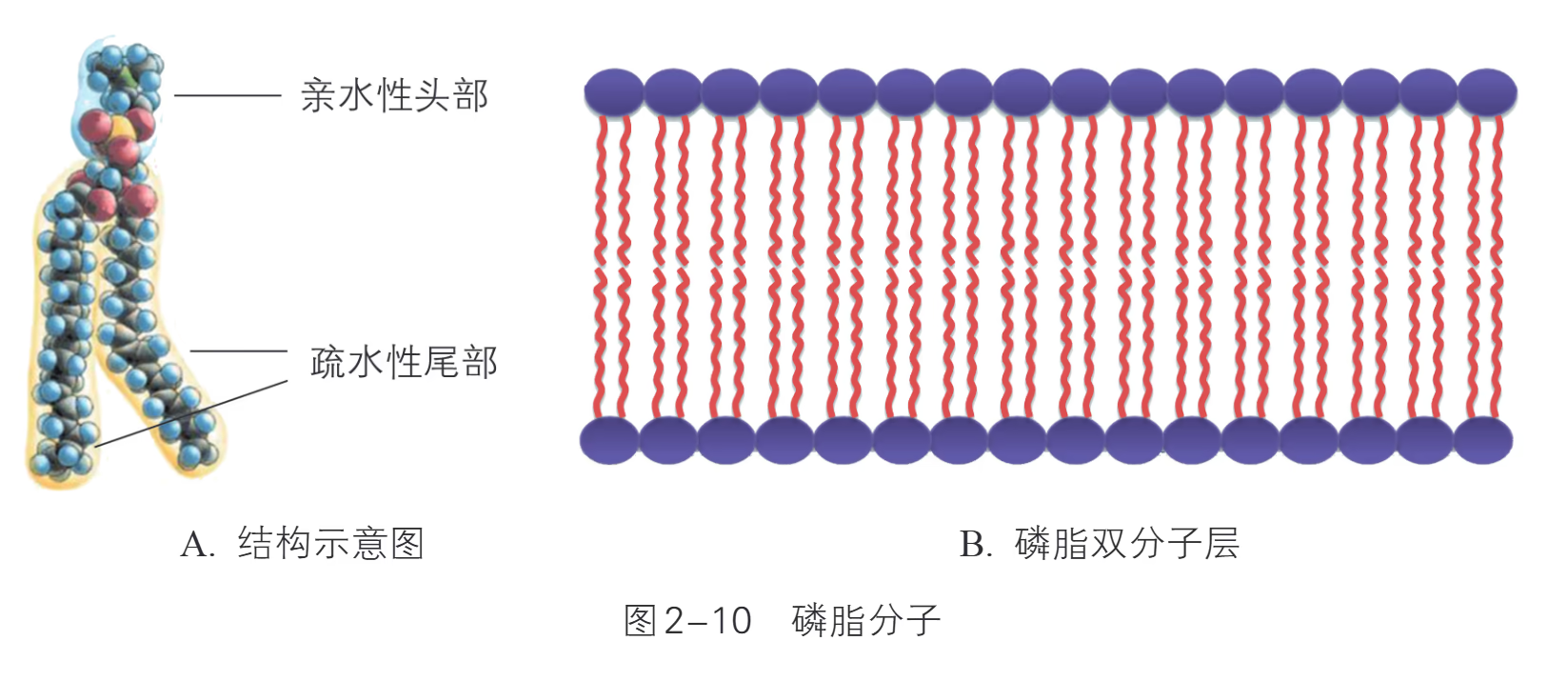
- 脂质(约 50%):磷脂是基本成分,其磷酸“头部”亲水,脂肪酸“尾部”疏水。在水中,磷脂分子自发形成磷脂双分子层,疏水端相对位于内侧,构成膜的基本支架。动物细胞膜还含有胆固醇,用于调节膜的流动性和稳定性。
- 蛋白质(约 40%):蛋白质是功能的主要执行者,其种类和数量决定了膜功能的复杂程度。膜蛋白以镶、嵌、贯穿等方式分布在脂双层中,担任转运蛋白、受体、酶或细胞连接的角色。
- 糖类(2%~10%):仅分布在细胞膜的外侧,与蛋白质结合形成糖蛋白(糖被),或与脂质结合形成糖脂。
【沪科技版】质膜上的蛋白质称为膜蛋白,有的覆盖在膜表面,有的镶嵌或贯穿在磷脂双分子层中。细胞的功能与膜蛋白种类和含量有关,有些细胞的膜蛋白仅占膜成分的 25%,有些则高达 75%。质膜中还有少量的糖,约 93% 与膜蛋白相连,形成糖蛋白;约 7% 与膜脂相连,组成糖脂。质膜内外两侧的分子并不对称,糖脂和糖蛋白上的糖均分布在质膜外侧,具有保护质膜和识别外界信息等功能。
目前公认的结构是 1972 年提出的流动镶嵌模型。
- 流动性:磷脂分子和大多数蛋白质分子都不是静止的,可以在膜平面内做横向移动,这使得膜能变形、融合或进行物质转运。
- 不对称性:膜内外两侧的分子种类和分布明显不同,例如糖链只位于外侧,这与细胞识别、信息接收等方向性功能有关。
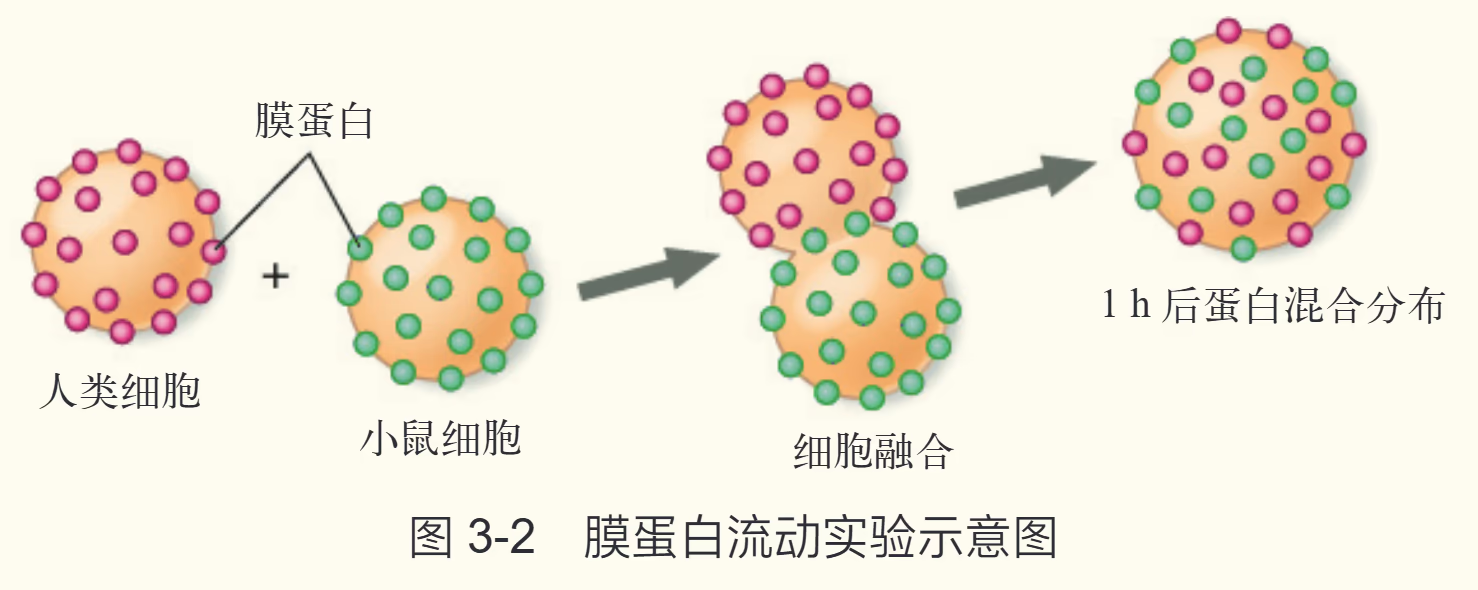
【浙科版】一些实验表明,生物膜并不是刚性的固态结构。1970 年,科学家进行了著名的人鼠细胞融合实验:先通过一些方法使人和小鼠的细胞融合成一个细胞,再用绿色荧光染料标记小鼠细胞表面的蛋白质分子,用红色荧光染料标记人细胞表面的蛋白质分子。开始时,融合的细胞一半呈现绿色,一半呈现红色。在 37 ℃下培养 40 min 后,两种颜色的荧光在整个细胞中都呈均匀分布。这表明细胞膜的蛋白质是呈流动状态的。
磷脂双分子层中间的疏水层通常情况下呈液态,磷脂分子在膜中的位置并不固定,使得磷脂分子及附着于其上的蛋白质分子发生运动。因此,生物膜具有一定的流动性。流动性是生物膜重要的结构特性,与生物膜进行能量转换、物质转运、信息传递和分裂等功能密切相关。
细胞膜在生命活动中承担着三项核心任务:
- 保障内部环境稳定:作为边界,它将细胞内物质与外界环境分隔开,避免胞内成分流失。
- 控制物质进出:膜具有选择透过性。
- 被动运输:顺浓度梯度,不消耗能量。包括自由扩散(如氧气、甘油)和协助扩散(需转运蛋白协助,如葡萄糖进入红细胞)。
- 主动运输:逆浓度梯度,消耗能量(ATP),需要载体蛋白,用于积累必需营养或排出废物。
- 胞吞与胞吐:针对大分子或颗粒物,通过膜的变形和囊泡转运完成。
- 信息交流:细胞通过分泌化学信号(如激素)、直接接触(如精卵识别)或胞间连丝/间隙连接来协调生命活动。
【浙科版】肥皂的主要成分与磷脂结构类似,是一端具有亲水性,另一端具有疏水性的两性分子。洗涤时,肥皂中两性分子的疏水端与油脂结合,将油脂包围起来,亲水端与周围的水分子结合,这样,被包裹的油脂在水中分散并形成乳浊液,再经过摩擦、振动,就被水冲洗掉了。这就是肥皂等去污剂去污的基本原理。
肥皂的去污原理是化学与物理作用共同的结果,其核心在于肥皂分子独特的两性(两亲性)结构。肥皂的主要成分是长链脂肪酸的盐(通常是钠盐或钾盐)。这种分子被称为**两性分子(Amphipathic molecule)**或两亲性分子,因为其结构中同时包含两个性质完全相反的区域:
- 亲水“头”部(Hydrophilic head):分子的一端是极性或带电荷的基团(如肥皂中的羧酸根离子)。由于水分子本身是极性分子,带有正电荷或负电荷的分子很容易与水结合,因此这一端极易溶于水。
- 疏水“尾”部(Hydrophobic tail):分子的另一端是长长的碳氢链(烃链)。这种结构是非极性的,不带电荷,无法与水分子形成良好的相互作用,因此具有憎水性。但它极易溶于油、脂等非极性有机溶剂。
这种“一头亲水,一头亲脂”的特性与生物膜中的磷脂分子结构非常相似。当我们使用肥皂清洗油渍时,去污过程通常分为以下几个物理化学阶段:
降低表面张力:肥皂属于表面活性剂(Surfactants)。当它溶解在水中时,肥皂分子会聚集在水面,亲水端指向水,疏水端指向空气,从而破坏水分子的氢键网络,降低水的表面张力。这使得水更容易润湿衣物或皮肤表面,渗透进污垢内部。
定向排列与包裹(微团形成):在洗涤过程中,肥皂分子的疏水端会避开水,主动与衣物或皮肤上的油脂结合并将油脂包围起来;而亲水端则朝向外侧与周围的水分子结合。随着肥皂浓度的增加,这些分子会形成一种球状聚集体,称为微团(Micelle):
- 微团内部:由无数个肥皂分子的疏水尾部组成,它们像一个“避风港”,将油垢卷入其中并紧紧锁住。
- 微团表面:布满了带电的亲水头部,确保整个聚集体能够稳定地悬浮在水中。
乳化与分散:通过手搓或洗衣机的揉搓、振动(机械作用),被肥皂分子包裹的油垢会从物体表面脱落,分解成极小的液滴分散在水中,这一过程称为乳化(Emulsification)。由于微团表面带有相同的电荷(如阴离子),它们之间会产生静电斥力,防止油滴重新聚集或再次沉积在物体表面。
机械移除与冲洗:最终,这些被包裹在微团中的油垢和随之粘附的微生物(细菌、病毒等)形成乳浊液。通过大量的清水冲洗,污垢随水流被机械性地带走,从而达到去污和**除菌(Degerming)**的目的。
特别补充:需要注意的是,普通的肥皂本身并不能直接杀死细菌或抑制其生长,因此它不被视为消毒剂或防腐剂。肥皂的作用主要是通过物理方式将细菌和病毒从皮肤表面“松动”并“搬运”走。除非肥皂中添加了特定的抑菌剂(如三氯卡班),否则其清洁效果主要依赖于机械性的冲洗过程。此外,肥皂在硬水(含有高浓度钙、镁离子的水)中容易形成难溶的沉淀(皂垢),降低洗涤效果,而合成去污剂(如 SDS)则能更好地适应硬水环境。
细胞膜的基本结构
对细胞膜成分的探索是生物学史上跨越数十年的经典研究历程,体现了“提出假说—实验验证—修正模型”的科学逻辑。以下是详细的阶段性总结:
脂质成分的初步推测(1895 年:欧文顿)
- 实验观察:欧文顿用 500 多种化学物质对植物细胞进行通透性实验,发现凡是溶于脂质的物质比不溶于脂质的物质更容易通过细胞膜。
- 推论:根据“相似相溶”原理,他推测细胞膜的主要组成成分中有脂质。
- 意义:这是人类首次通过间接推理对膜成分做出的定性判断。
磷脂双分子层的确立(1925 年:戈特和格伦德尔)
- 实验操作:他们用丙酮从人的红细胞中提取脂质,并在空气—水界面上铺成单分子层。
- 关键发现:测得单分子层的面积恰好是红细胞表面积的 2 倍。
- 结论:细胞膜中的磷脂分子必然排列为连续的两层。
- 原因:磷脂分子具有亲水的“头”和疏水的“尾”。在细胞内外均为水环境的条件下,疏水尾部相对排列在内侧,亲水头部向着两侧,从而自发形成稳定的双分子层结构。
蛋白质成分的加入与“三明治”模型
- 张力实验(1935 年):丹尼利和戴维森发现细胞膜的表面张力远低于油—水界面。由于油脂滴表面吸附蛋白质后张力会降低,他们推测膜中附有蛋白质。
- 电镜观察(1959 年:罗伯特森):利用电子显微镜观察,发现质膜呈现“暗—亮—暗”三条带。
- 静态模型:他提出“单位膜模型”,认为蛋白质分布在脂质双分子层的两侧,像三明治一样。他将此描述为静态的统一结构,但这无法解释细胞生长和变形运动。
膜的流动性验证(1970 年,人—鼠细胞融合实验)
- 实验方法:用荧光染料标记人、鼠细胞表面的蛋白质,诱导融合。
- 现象:37 ℃环境下培养 40 min 后,两种颜色的荧光均匀分布。
- 意义:该实验有力地证明了细胞膜具有流动性,否定了静态模型的说法。
流动镶嵌模型的完善(1972 年:辛格和尼科尔森)
- 基本支架:磷脂双分子层。
- 蛋白质排布:以覆盖、镶嵌、贯穿等方式分布,且分布是不对称的。
- 运动特性:磷脂分子和大多数蛋白质分子都能运动。
- 糖类分布:外表面含有糖蛋白(糖被)或糖脂,负责细胞识别和信息传递。
【沪科技版】质膜的基本骨架是磷脂双分子层。磷脂分子亲水性的头部朝向细胞内外两侧,疏水性的尾部相对,在磷脂双分子层内部形成一层疏水的屏障。紧密排列的磷脂分子不允许大分子物质进出,疏水屏障将细胞内外的极性小分子和离子等物质隔开,从而将细胞与生活环境分开,使细胞内部形成相对稳定的环境。临床上应用这个原理可以判断细胞是否有损伤。例如,正常情况下谷丙转氨酶(ALT)存在于人体肝细胞中,而血清中含量极少。当肝细胞质膜受损时,ALT 会流出细胞而进入血液。因此,血清中 ALT 量的变化可作为临床上诊断肝功能的指标之一。
对于植物细胞来说,细胞壁是全透性的,一般提供支持和保护作用,其细胞系统边界是细胞膜,而非细胞壁。
流动镶嵌模型(fluid mosaic model)认为,细胞膜主要是由磷脂分子和蛋白质分子构成的。磷脂双分子层是膜的基本支架,其内部是磷脂分子的疏水端,水溶性分子或离子不能自由通过,因此具有屏障作用。蛋白质分子以不同方式镶嵌在磷脂双分子层中:有的镶在磷脂双分子层表面,有的部分或全部嵌入磷脂双分子层中,有的贯穿于整个磷脂双分子层。这些蛋白质分子在物质运输等方面具有重要作用。
细胞膜不是静止不动的,而是具有流动性,主要表现为构成膜的磷脂分子可以侧向自由移动,膜中的蛋白质大多也能运动。细胞膜的流动性对于细胞完成物质运输、生长、分裂、运动等功能都是非常重要的。
对细胞膜的深入研究发现,细胞膜的外表面还有糖类分子,它和蛋白质分子结合形成糖蛋白,或与脂质结合形成糖脂,这些糖类分子叫作糖被(glycocalyx)。糖被在细胞生命活动中具有重要的功能。例如,糖被与细胞表面的识别、细胞间的信息传递等功能有密切关系。
【浙科版】细胞膜上的蛋白质统称为膜蛋白。膜蛋白也和磷脂分子一样,有水溶性部分和脂溶性部分。通常细胞的功能越多,其膜蛋白的种类和数量就越多。有些膜蛋白选择性地吸收或排出特定物质,控制其进出细胞,这类膜蛋白称为转运蛋白。例如,细胞吸收离子、葡萄糖、氨基酸等营养物质都需要通过特定的转运蛋白来完成。有些膜蛋白催化特定化学反应,例如,小肠上皮细胞的某些膜蛋白可促进食物中营养物质的消化分解。同一组织的相邻细胞间通过某些膜蛋白紧密连接在一起。某些膜蛋白接收外界信息或刺激,激活细胞内相应生理生化反应,在细胞与外界的信息交流和细胞识别中发挥重要作用,如胰岛素受体。细胞膜的功能主要通过膜蛋白来实现。
细胞外被(cell coat)又称糖萼(glycocalyx),存在于细胞膜外表面,由多糖与细胞膜中的蛋白质或脂质结合而成,是细胞膜的正常成分。细胞外被不仅对细胞膜起保护作用,而且在细胞识别过程中起重要作用。最早关于细胞识别的观察,是将两种不同的海绵动物的细胞分散成单个细胞,然后混合培养。结果发现,只有同种海绵的细胞才能结合在一起。这说明细胞能相互识别。通过对细胞表面成分的分析,发现两种海绵细胞表面的糖蛋白有差异,其差异主要表现为糖类的不同。
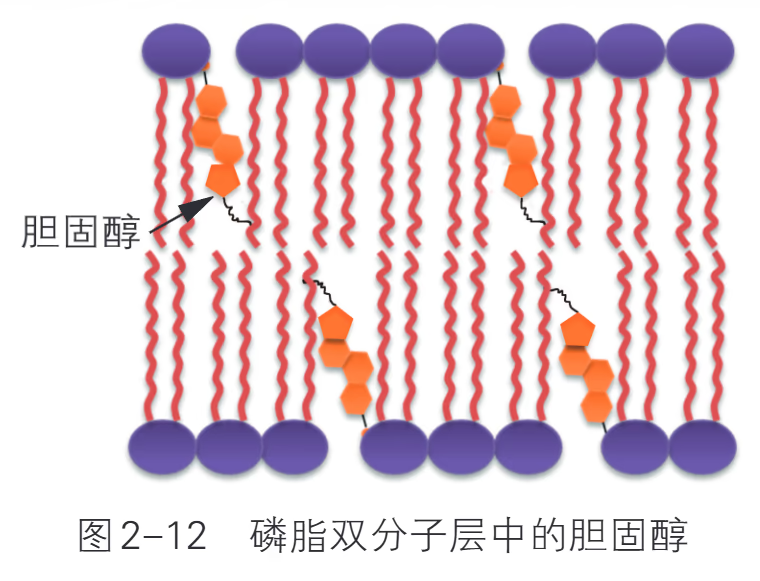
胆固醇存在于动物细胞膜中,占细胞膜所有脂质的 10%~30%。植物细胞膜一般不含胆固醇,但含有其他固醇类化合物。在动物细胞中,胆固醇能保持细胞膜的稳定性。胆固醇分子主要位于磷脂双分子层的疏水环境中,具有一个很小的亲水基团,与磷脂头部连接。胆固醇对细胞膜中磷脂分子的活动具有双重调节作用。一方面,胆固醇通过与磷脂脂肪酸链的相互作用,具有限制其运动、增加其有序性的作用;另一方面,胆固醇也能将磷脂分子分隔开以增强其运动性,保证细胞膜在通常情况下处于流动状态。这种双重的调节作用取决于环境和胆固醇的数量。例如,环境温度较低时,胆固醇更有助于保持细胞膜的柔韧性;细胞膜外侧磷脂分子中胆固醇的含量往往高于内侧,因此内侧的磷脂活动性较弱。
除细胞膜外,细胞内还存在其他的膜结构,它们构成了细胞核等多种结构的边界。细胞内所有的膜称为生物膜(biomembrane)。虽然细胞内不同部位的膜功能有所不同,但它们都具有相似的物质组成和空间结构。
在细胞生物学中,细胞质膜)的“流动镶嵌模型”不仅描述了膜的流动性,更强调了膜在空间上的不对称性(Asymmetry)与功能上的区域化(Regionalization/Membrane Domains)。这种精密的组织结构是细胞完成复杂生命活动的物质基础。细胞膜的不对称性 (Membrane Asymmetry):不对称性是指细胞膜内外两层的成分、结构和功能存在显著差异。这种差异在膜合成之初便已确定,并贯穿其生命周期。
脂质分布的不对称性
- 种类差异:脂双层的内外两层所含的磷脂种类不同。例如,在动物细胞中,磷脂酰胆碱(PC)和鞘磷脂常集中在膜的外侧(非胞质面),而磷脂酰乙醇胺(PE)和磷脂酰丝氨酸(PS)则主要分布在内侧(胞质面)。
- 形成机制:不对称性起始于内质网和高尔基体。内质网中的**磷脂转位蛋白(Scramblase)负责随机平衡脂质,而高尔基体中的翻转酶(Flippase)**则选择性地将特定磷脂从外侧翻转至内侧,从而建立了这种不对称分布。
- 功能意义:特定脂质的分布与信号转导有关,例如磷脂酰肌醇(PI)主要分布在内侧以传递胞内信号。
膜蛋白分布与定向的不对称性
- 固定取向:每种跨膜蛋白在脂双层中都有特定的方向(Orientation),这决定了其功能域的朝向。例如,受体的配体结合位点必须面向细胞外,而信号传导区则面向胞质溶胶。
- 合成决定:这种定向在蛋白质合成并插入内质网膜时就已确定,且在随后的囊泡转运和膜融合过程中始终保持不变。
糖类的绝对不对称性
- 分布规律:糖蛋白和糖脂上的糖链仅分布在质膜的外表面,形成糖被(糖萼)。
- 功能:糖被负责细胞识别、信息交流、保护和润滑细胞表面。
细胞膜的区域化 (Membrane Regionalization) 是指细胞通过特定机制限制膜成分(尤其是蛋白质)的随机扩散,使其集中在特定的膜区(膜域),以高效执行功能。
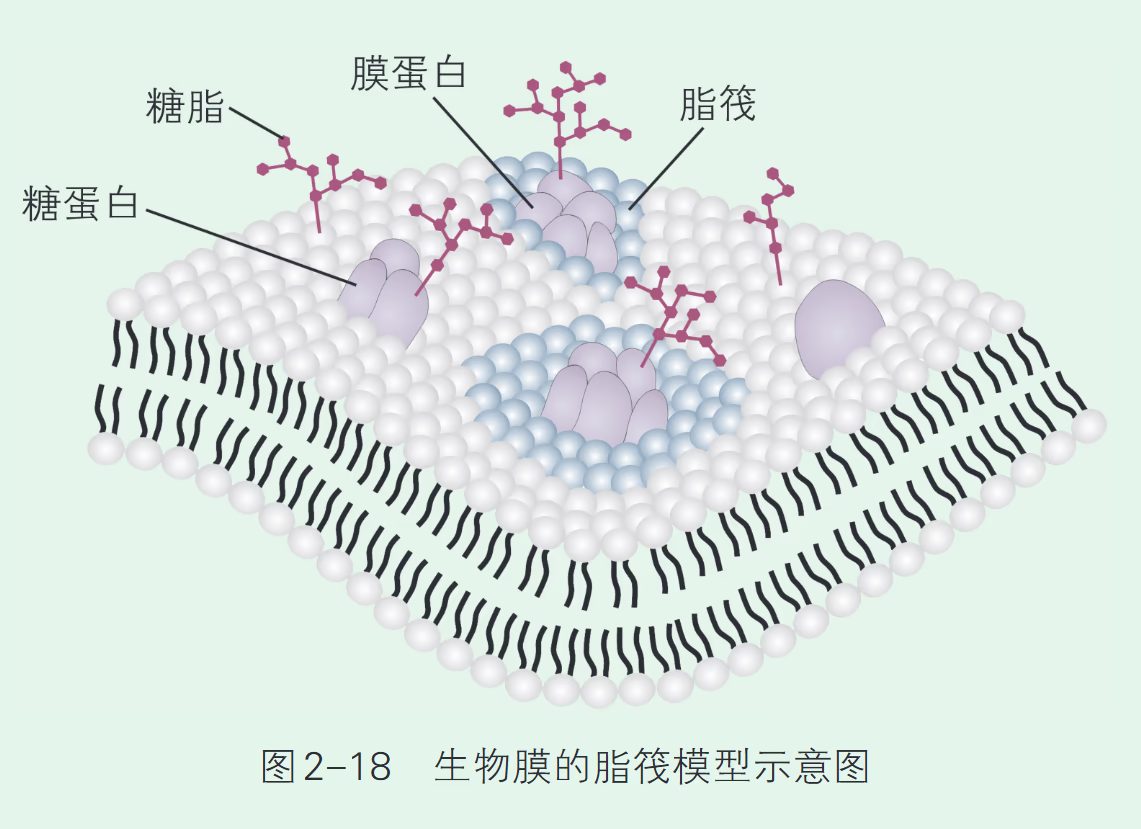
【浙科版】20 世纪 80 年代,有人提出生物膜有许多胆固醇聚集的微结构区,就像水面上漂浮的竹筏一样,称为脂筏模型。脂筏位于生物膜的外侧,胆固醇与磷脂相互作用,形成了脂筏的基本结构。一些膜蛋白与脂筏表面的化学基团结合,被“锚定”在一起,构成了生物膜上分子排列紧密、结构相对稳定的特定区域。所以,脂筏像个蛋白质停泊平台一样,有助于蛋白质执行相应功能。这一模型可以解释生物膜的某些性质和功能,但仍需要更多的证据支持。
膜域 (Membrane Domains) 的概念:细胞并非将所有蛋白质均匀混合在“脂海”中,而是将具有相似功能的蛋白质聚集在特定的区域。
为了防止膜蛋白因流动性而均匀散开,细胞采用了以下锚定和屏障机制:
- 胞内锚定(细胞骨架限制):膜蛋白通过附着蛋白与细胞皮层(如红细胞的血影蛋白网)相连,使其相对固定。
- 胞外锚定(细胞外基质限制):膜蛋白被连接到细胞外的纤维结构(如细胞外基质分子)上。
- 相邻细胞锁定:某些蛋白质通过与相邻细胞表面的特定蛋白质结合而被固定在接触位点。
- 物理屏障(扩散屏障)紧密连接 (Tight Junctions):在肠上皮细胞等组织中,紧密连接像一道“篱笆”,阻断了膜蛋白在顶端膜和基底侧膜之间的侧向扩散,维持了上皮细胞的极性。
功能微区:脂筏模型 (Lipid Raft Model)。
- 结构特征:脂筏是膜上富含胆固醇、鞘磷脂和特定膜蛋白的微小区域。
- 功能特点:这些区域结构致密、流动性较低,像漂浮的“木筏”一样募集特定的信号分子。脂筏作为蛋白质停泊平台,极大地提高了信号转导、跨膜运输和病原体侵染过程的效率。
流动性与有序性的平衡:
- 流动性是前提:允许蛋白质相互碰撞、结合并执行任务。
- 不对称性是基础:确立了细胞内外环境的物理和化学边界。
- 区域化是手段:通过锚定和屏障机制,使细胞膜从“无序的海洋”变为“高度组织化的加工厂”。
理解要点:细胞膜具有流动性,但并不意味着它是杂乱无章的。由于不对称性和区域化的存在,质膜呈现出高度精密的、空间分布不均一的特征。
细胞膜酶蛋白:在细胞生命活动中,细胞质膜不仅仅是一层简单的屏障,它更是一个高度动态的、功能复杂的生化反应平台。在这个平台上,镶嵌、嵌入或贯穿于磷脂双分子层中的 “酶”蛋白(Membrane-bound Enzymes) 发挥着至关重要的作用。以下根据提供的来源,从结构特征、功能分类、工作机制及其在生物学中的意义四个维度详细展开讲解细胞膜上的酶蛋白。
细胞膜上的酶绝大多数属于蛋白质,少数为具有催化功能的 RNA(核酶)。它们在膜上的分布表现出明显的不对称性和流动性。
- 膜关联方式:
- 膜内在蛋白(整合蛋白):许多酶蛋白整个地穿过脂双层,其疏水区(通常为 \alpha 螺旋)位于膜内部,亲水区则暴露在膜两侧。例如,腺苷酸环化酶含有 12 段穿膜螺旋。
- 膜周边蛋白:某些酶并不嵌入脂双层,而是通过非共价相互作用附着在膜内在蛋白或脂质头部。
- 脂质锚定蛋白:部分酶通过共价连接的脂质分子(如糖基磷脂酰肌醇 GPI)锚定在膜表面。
- 流动性:构成膜的酶蛋白大多可以侧向自由移动,这种流动性对于它们寻找底物或与其他信号分子碰撞至关重要。
- 微区定位(脂筏):某些酶(如特定的受体激酶)并非均匀分布,而是聚集在富含胆固醇和鞘磷脂的“脂筏”微区内,形成功能性的信号转导平台。
膜酶根据其催化的反应类型,可以分为以下几大核心类别:
信号转导类酶(核心通讯员):这类酶是细胞响应外界刺激的关键,负责将细胞外的化学信号转变为细胞内的生理反应。
腺苷酸环化酶 (AC):催化 ATP 去掉焦磷酸生成第二信使 cAMP。它是 G 蛋白偶联受体通路的下游核心效应器。
磷脂酶 C (PLC):能断裂磷脂(如 PIP2)释放出两个重要的第二信使:三磷酸肌醇 (IP3) 和二酰基甘油 (DAG)。
蛋白激酶 (Kinases):膜上的酶联受体(如受体酪氨酸激酶 RTK)在结合配体后,其胞内域活化,催化自身或底物蛋白磷酸化,引发信号级联反应。
蛋白磷酸酶:与激酶相对应,负责水解蛋白质上的磷酸基团,起到关闭信号通路的作用。
能量转换与离子转运类酶(分子泵):许多被称为“泵”的穿膜蛋白实质上是具有ATPase 活性的酶,它们利用 ATP 水解提供的能量进行主动运输。
Na^+/K^+-交换 ATP 酶(钠钾泵):每消耗一个 ATP,逆电化学梯度泵出 3 个Na^+并泵入 2 个K^+。它既是转运蛋白,也是催化 ATP 水解的酶。
Ca^{2+}-ATP 酶(钙泵):位于质膜或内质网膜上,负责将细胞质中的Ca^{2+}泵出或泵入细胞库,维持胞质低钙状态。
代谢与消化类酶(边界加工厂):
刷状缘酶:位于小肠上皮细胞微绒毛膜上的酶(如蔗糖酶、乳糖酶、肽酶),负责将食物残渣进一步水解成可吸收的小分子。
分泌型膜蛋白酶:如基质金属蛋白酶 (MMP),可以降解细胞外基质,参与组织重塑和肿瘤迁移。
合成与标志类酶:
纤维素合成酶:位于植物细胞膜上,负责合成细胞壁的纤维素成分。
糖基转移酶:参与质膜表面糖被(糖蛋白、糖脂)的构建,这些糖链对于细胞识别至关重要。
膜酶的工作机制与特性:
- 降低活化能:膜酶通过与底物形成特定的酶 - 底物复合物,降低反应所需的活化能,使生化反应在常温常压的温和条件下快速进行。
- 专一性(锁钥学说):每种膜酶的活性部位具有特定的空间结构,只能与特定形状的底物分子契合。例如,蔗糖酶只能催化蔗糖水解,而不能催化麦芽糖。
- 高效性:膜酶的催化效率远高于无机催化剂(如Fe^{3+}),通常高出10^7至10^{13}倍。
- 反应环境要求:膜酶活性受温度和 pH 值的显著影响。
- 最适条件:人体膜酶的最适温度通常在 35-40℃,pH 多接近中性。
- 失活:强酸、强碱或高温会破坏酶的空间结构,导致永久失活。
临床应用与生物学意义:
- 疾病诊断指标:
- 正常情况下,某些酶(如谷丙转氨酶 ALT)存在于肝细胞内,当细胞膜受损或透性改变时,这些酶会大量释放入血,成为肝功能受损的指标。
- 佝偻病患者血清中的碱性磷酸酶活性会增高。
- 药物靶点:
- 许多药物通过调节或抑制膜酶活性起作用。例如,他汀类药物通过抑制肝脏中合成胆固醇的关键酶(HMG-CoA 还原酶)来降低血脂。
- 许多杀虫剂(如有机磷农药)通过抑制神经突触膜上的胆碱酯酶活性导致昆虫中毒死亡。
- 生命活动的有序化:膜酶在细胞内的分区定位(区域化),使得各种不相容的生化反应(如合成与分解)能同时且互不干扰地进行,极大地提高了代谢效率。
综上所述,细胞膜上的酶蛋白是细胞代谢、能量转换和信号交流的核心执行分子,其精细的结构与定位直接决定了细胞生命活动的复杂性与高效性。
【沪科技版】虽然磷脂双分子层对细胞内外的物质进出有屏蔽作用,但实际上,细胞内外的物质可以通过质膜进行有选择性的交换。其中,膜蛋白在物质交换中起着重要作用。
生物体内的细胞并不是孤立存在的,就像人与人之间要进行信息交流一样,细胞之间也需要通过信息交流才能相互协调,共同完成生理活动。例如,神经细胞通过分泌神经递质作用于靶细胞,内分泌细胞则通过分泌激素作用于靶细胞。有些称为受体的膜蛋白,可用于接受神经递质、激素等特定信号分子。
受体接受信号分子后,激发细胞内特定的生命活动,对信号作出应答。若信号传递受阻,则可能导致疾病。例如,胰岛β 细胞分泌的胰岛素通过血液循环到达肝细胞,被肝细胞质膜上的胰岛素受体识别,可促进肝细胞吸收葡萄糖,从而降低血糖浓度。如果胰岛素受体结构发生改变,不能识别胰岛素,影响肝细胞对葡萄糖的吸收,会导致血糖控制失调,可能会引发Ⅱ 型糖尿病。
不同种类的细胞质膜表面的糖蛋白、糖脂的种类不同,具有特异性。细胞可通过糖蛋白和糖脂等进行细胞之间的信息识别。例如,在炎症反应过程中,中性粒细胞(一种白细胞)在炎症部位的聚集过程与此相关。在细菌感染初期,感染处的血管内皮细胞在质膜表面形成特殊的糖蛋白(称为凝集素),可被白细胞表面的糖蛋白和糖脂特异性识别,引起白细胞在此处聚集并穿过血管壁进入感染部位,从而杀灭细菌。
【沪科技版】随着社会和科技的发展,应用靶向技术向肿瘤区域精确递送药物的“靶向治疗”和控制肿瘤生长或代谢的“靶点治疗”成为肿瘤研究的热点。根据靶向部位的不同,又可以将肿瘤靶向治疗分为两大类,即肿瘤细胞靶向治疗和肿瘤血管靶向治疗。肿瘤细胞的靶向治疗能使药物更特异性地针对目标,在提高药物治疗效果的同时减少了对正常细胞的毒副作用。如果我们能更多地了解肿瘤细胞的结构特征,发现更多的特异性靶点,例如膜蛋白、细胞增殖相关因子等,便能研发出更多的治疗药物,加快解决肿瘤危害人类健康的难题。
肿瘤细胞靶向治疗是利用肿瘤细胞表面的特异性受体作为靶向,药物与肿瘤细胞特异性结合,抑制肿瘤细胞生长增殖,促使其死亡。例如,细胞表面的表皮生长因子受体(EGFR)接受信号后介导细胞的生长、增殖和分化等生理过程。与正常细胞相比,多种肿瘤细胞表面的 EGFR 数量极高。针对 EGFR 靶向的治疗药物(如小分子化合物、单克隆抗体等)能阻断 EGFR 与信号分子结合,从而抑制肿瘤细胞的增殖并诱导其死亡。目前 EGFR 的靶向药物在肺癌、结直肠癌等治疗中广泛应用。
肿瘤血管靶向治疗则是利用肿瘤区域新生毛细血管内皮细胞表面的特异性受体起作用。针对肿瘤细胞的药物要到达肿瘤细胞靶区,需要通过血管内皮细胞屏障进入肿瘤组织环境,这一过程是相对缓慢的。针对血管靶向的药物则有很大的优势,在给药后只需积聚在血管靶标部位,抑制肿瘤组织的血管生成,阻断机体通过血液对肿瘤组织提供营养物质,以达到“饿死”肿瘤细胞的目的。同时也阻断肿瘤细胞通过血管进行转移的途径。血管表面的血管内皮生长因子受体(VEGFR)在特异性信号分子血管内皮生长因子(VEGF)的刺激下,会促进血管内皮细胞增殖和新生血管的形成。在针对恶性肿瘤转移的治疗中,多采用针对 VEGF/VEGFR 的靶向药物,以阻断肿瘤细胞通过血管转移。
细胞器之间的分工
细胞是生命活动的基本单位,其内部由多种细胞器分工合作,共同维持生命运转。以下是各主要细胞器的详细结构与功能:
双层膜结构:能量转换站
线粒体 (Mitochondrion):由内外两层膜构成,内膜向内腔折叠形成嵴,大大增加了内膜面积;基质中含有少量 DNA、RNA 和核糖体。细胞进行有氧呼吸的主要场所,为生命活动提供能量(约占 95%),被称为“动力车间”。
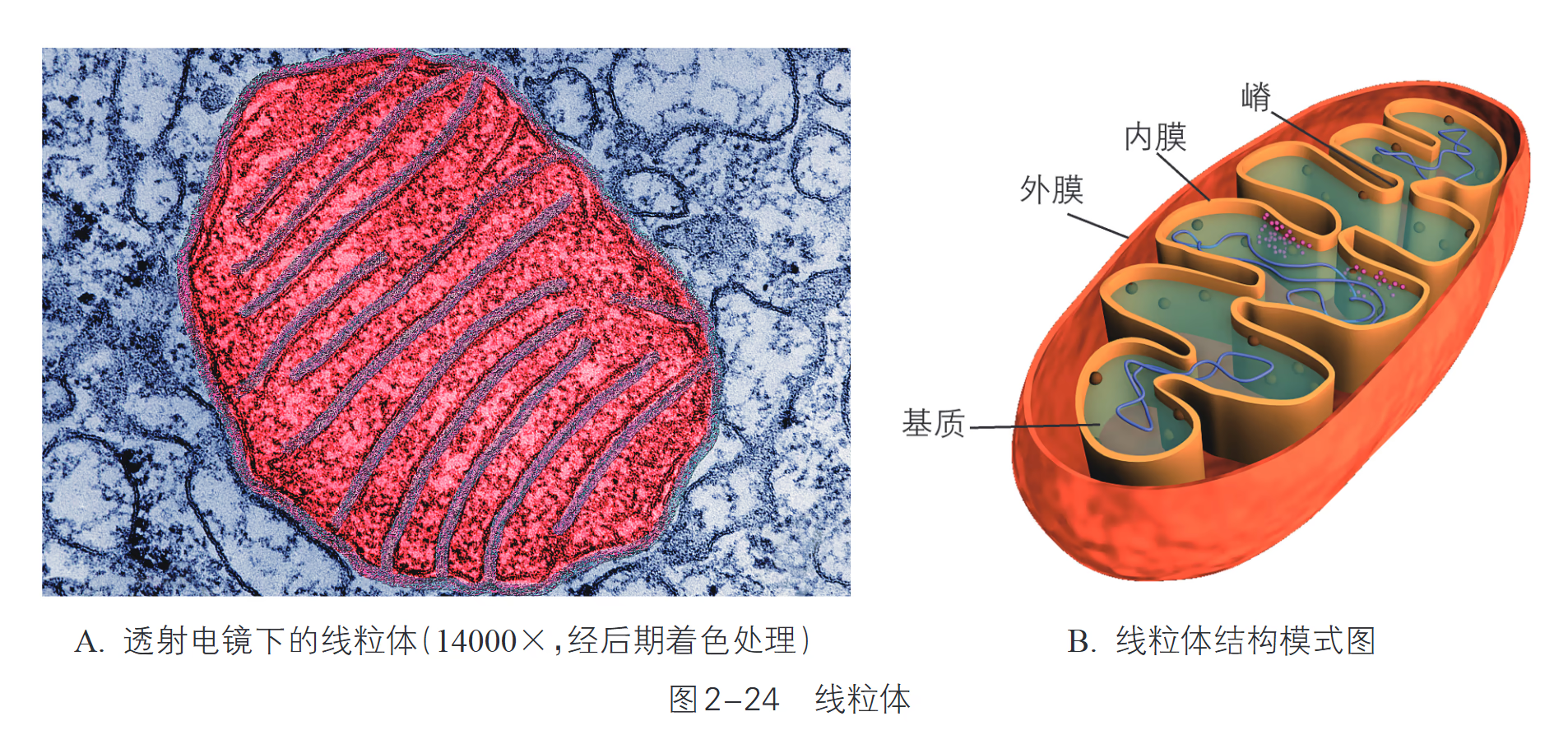
【浙科版】线粒体(mitochondria)是真核细胞非常重要的细胞器,一般呈颗粒状或短杆状,相当于一个细菌的大小,由内、外两层膜构成。外膜平整,内膜向内凹陷形成嵴(cristae)。嵴的形成有助于增大内膜的表面积,有利于生化反应的进行。内、外膜之间及内部是液态的基质,富含多种酶。线粒体基质中具有 DNA、RNA 和核糖体,能合成一部分自身所需的蛋白质。
叶绿体 (Chloroplast):由内外两层膜、类囊体和基质组成;类囊体堆叠形成基粒,膜上分布着光合色素和酶。绿色植物进行光合作用的场所,是“养料制造车间”和“能量转换站”。

【浙科版】叶绿体(chloroplast)是存在于植物细胞中进行光合作用的细胞器,呈球形或椭球形,具有双层膜结构,内部是液态的基质。基质中具有复杂的膜结构,称为类囊体,与光合作用有关的色素附着于这些膜上。类囊体可堆叠成基粒。叶绿体基质中也具有 DNA、RNA 和核糖体。关于叶绿体和光合作用的具体内容在第三章第五节中详细介绍。
单层膜结构:合成与加工车间
内质网 (Endoplasmic Reticulum):由膜围成的管状、泡状或扁平囊状结构连接而成的连续管道系统,因为最初是在细胞质内部发现的网状结构得名。分类与功能:粗面内质网(附着核糖体)参与蛋白质的合成、加工和运输;光面内质网是脂质合成的场所,还具有解毒等功能。

【浙科版】科学家用电子显微镜观察小鼠细胞时,发现细胞质内具有极其发达的网状结构,称之为内质网(endoplasmic reticulum,ER)。内质网是由一系列片状的膜囊和管状的腔组成,向内连接细胞核的核膜,向外连接细胞膜,并与高尔基体相互联系,构成了细胞内庞大的物质运输通道。内质网有两种类型,即光面内质网(smooth ER)和粗面内质网(rough ER)。光面内质网常为管状,是运输蛋白质和合成脂质的重要场所。构成生物膜的磷脂和胆固醇几乎全部由光面内质网合成。另外,某些细胞的光面内质网非常发达并具有特殊功能。例如,肝细胞光面内质网有解毒功能,一些内分泌细胞的光面内质网可以合成类固醇激素。粗面内质网多呈扁囊状,因表面附着许多小颗粒而得名。这些小颗粒称为核糖体(ribosome),由核糖体合成的蛋白质进入内质网,进一步被加工和运输。
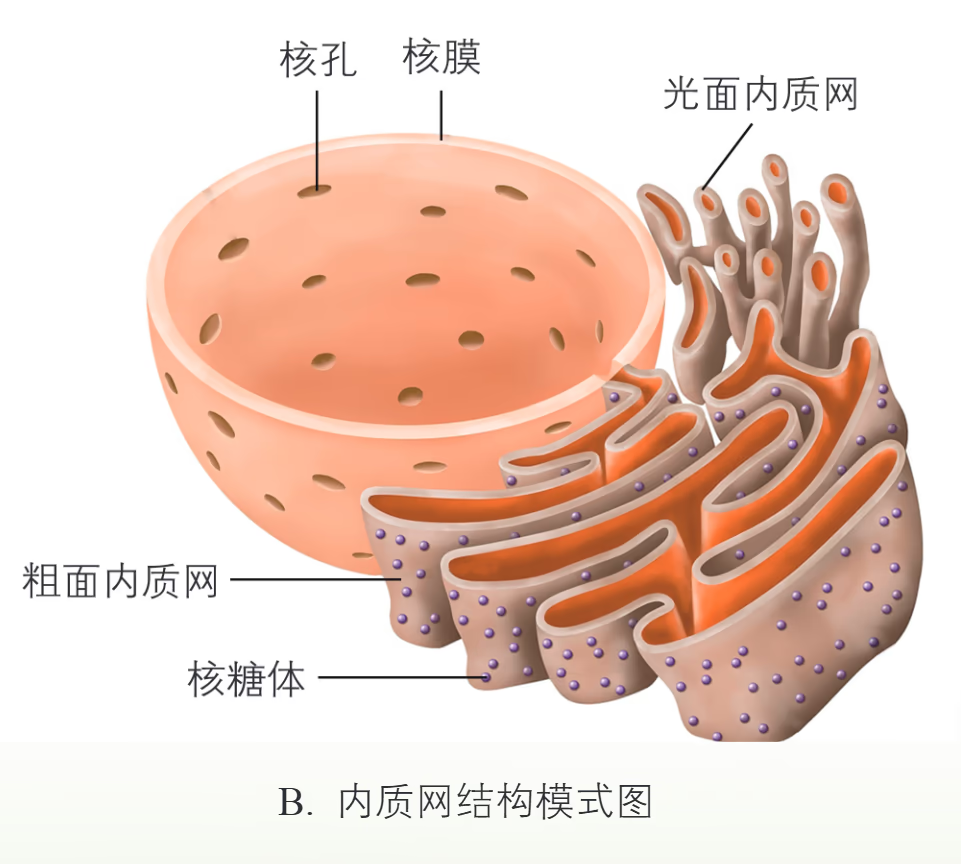
高尔基体 (Golgi Apparatus):最初由意大利医生高尔基发现,又称做高尔基复合体。由扁平膜囊和大小不一的囊泡堆叠而成。对来自内质网的蛋白质进行加工、分类和包装的“车间”;在植物细胞中还参与细胞壁的形成。
溶酶体 (Lysosome):单层膜构成的囊泡,内部含有 60 种以上的水解酶。细胞的“消化车间”,能分解衰老损伤的细胞器,吞噬并杀死侵入细胞的病毒或细菌。
【浙科版】溶酶体的主要功能是进行细胞内消化,它能消化细胞从外界吞入的颗粒、自身衰老的细胞器和碎片。细胞从外界吞入物质后形成吞噬泡,吞噬泡与溶酶体融合,其中的水解酶便将吞噬泡中的物质降解。溶酶体可将消化作用局限在特定结构中,这对保证细胞中其他结构的完整性具有重要意义。
液泡 (Vacuole):主要存在于植物细胞中,内含细胞液(含无机盐、糖类、色素等)。调节植物细胞内的环境,充盈时保持细胞坚挺。
【浙科版】液泡(vacuole)是由单层膜构成的充满水溶液的泡状细胞器,主要存在于植物细胞中。植物根尖、茎尖等部位刚分裂形成的细胞中,液泡很小,但数量很多。随着细胞逐渐成熟,小液泡融合成大液泡,并占据细胞中央,细胞质和细胞核都被挤到四周,紧贴着细胞膜。具有中央大液泡是成熟植物细胞的显著特征。
植物细胞液泡中的水溶液称为细胞液,含有无机盐、糖类、氨基酸、色素等。液泡中的色素使某些植物的花、叶、果实呈现不同的颜色。液泡中的细胞液为植物细胞储存水分和营养物质,调节细胞渗透压平衡、酸碱平衡、离子平衡,维持细胞正常形态。液泡还富含水解酶,能吞噬衰老的细胞器,其作用与动物细胞的溶酶体相似。所以,液泡对维持植物细胞正常生命活动具有重要的意义。
无膜结构:蛋白质与分裂助手
核糖体 (Ribosome):由 RNA 和蛋白质组成,无膜结构。合成蛋白质的场所,是“生产蛋白质的机器”。

【浙科版】核糖体是一种无膜的细胞器,由 RNA 和蛋白质构成,是合成蛋白质的场所。核糖体有两种存在形式,合成的蛋白质功能有所不同:游离于细胞溶胶的核糖体合成的蛋白质,通常用于细胞自身或构成自身结构,如红细胞中的血红蛋白、肌细胞的肌纤维蛋白等;附着于粗面内质网等结构的核糖体合成的蛋白质,被运输至胞外或细胞的其他部位。核糖体在细胞中的位置并不是固定不变的,两种存在形式可以相互转化。
中心体 (Centrosome):分布在动物和低等植物细胞中,由两个互相垂直的中心粒及周围物质组成。与细胞的有丝分裂有关。
支持系统:细胞骨架与细胞壁
细胞骨架 (Cytoskeleton):由蛋白质纤维(微管、微丝、中间丝)组成的网架结构。维持细胞形态,锚定并支撑细胞器,参与细胞运动、分裂、分化及物质运输。
细胞壁 (Cell Wall):位于植物质膜外,主要成分是纤维素和果胶,具有全透性。
对细胞起支持与保护作用。
**细胞溶胶(cytosol)**又称细胞质基质(cytoplasmic matrix),是细胞内除去细胞器以外的胶状物质。通常情况下,细胞溶胶的含水量占细胞总体积的 70\% 左右。细胞溶胶含有丰富的蛋白质,还含有糖类、氨基酸、无机盐等多种营养物质,是细胞与外界环境、细胞质与细胞核及细胞器之间物质运输、能量交换和信息传递的重要介质。
细胞溶胶也是许多代谢反应的重要场所。例如,真核生物的需氧呼吸就是在细胞溶胶和线粒体内进行的;细胞内的许多蛋白质都是在细胞溶胶中游离核糖体上合成,继而分别转运到细胞核、叶绿体、线粒体等结构中,也有些蛋白质成为细胞溶胶中一些结构的组成成分。此外,细胞溶胶还参与某些脂质的合成、蛋白质的加工和降解、大分子物质和细胞器的移动等。细胞溶胶所承担的功能往往不是孤立单一的,而是与细胞内其他结构相互配合,协同完成各项生命活动。
细胞内部这些精巧的结构并非孤立存在,而是通过生物膜系统在结构和功能上紧密联系。

细胞骨架(Cytoskeleton)是真核细胞细胞质中由蛋白质纤维组成的精密网架结构。它并非如人类骨骼般固定不变,而是一个高度动态的系统,在细胞改变形状、分裂及应对环境挑战时会不断重组。细胞骨架主要由三种类型的蛋白质纤维构成,它们在结构、成分和功能上各具特色:
- 微管 (Microtubules)
- 结构与成分:微管是细长的中空管状结构,外径约 25nm。它由微管蛋白(Tubulin,包括 \alpha 和 \beta 两类亚基形成的二聚体)聚合而成。
- 特性:具有极性,且表现出“动态不稳定性”,即在生长期和缩短期之间快速转换。
- 功能:
- 胞内运输:作为囊泡、细胞器和生物大分子移动的“轨道”。
- 细胞分裂:在有丝分裂中形成纺锤体,负责将染色体精确地拉向两极。
- 运动器官:是中心体、纤毛和鞭毛的核心结构成分。
- 微丝 (Microfilaments / Actin Filaments)
- 结构与成分:直径约 7nm,是由球状肌动蛋白(Actin)单体相互缠绕形成的实心双螺旋纤维。
- 特性:柔韧且富有弹性,主要集中在细胞质膜内侧的细胞皮层(Cell Cortex)中。
- 功能:
- 维持与改变形状:产生张力以支持细胞形状,并驱动细胞表面的运动。
- 运动机制:参与肌肉收缩、变形虫运动(伪足形成)以及植物细胞中的胞质环流。
- 胞质分裂:在动物细胞分裂末期形成收缩环,将细胞缢裂为二。
- 中间纤维 (Intermediate Filaments)
- 结构与成分:直径约 8-12nm(通常取 10nm),介于微管和微丝之间。它由多种纤维蛋白(如角蛋白、波形蛋白、核纤层蛋白)装配成绳索状。
- 特性:在三类纤维中最坚韧、最耐久,具有极强的抗拉强度,不易解聚。
- 功能:
- 机械支撑:增强细胞承受机械压力的能力,防止细胞受压破裂。
- 固定位置:将细胞核等细胞器固定在特定位置,并在核被膜下形成核纤层以稳定细胞核。
细胞骨架不仅仅是细胞的“骨骼”(支撑),也是它的“肌肉”(动力系统)和“高铁运输线”。
- 维持形态与内部有序性:它赋予细胞特定的形状,并锚定支撑许多细胞器,保持细胞内部结构的有序性。
- 物质运输与能量转换:细胞内的各类小泡和细胞器沿着细胞骨架定向转运。例如,驱动蛋白和动力蛋白(马达蛋白)会携带“货物”在微管上“行走”。
- 细胞运动:无论是白细胞的迁移、精子的游动,还是细胞内部的物质流动,都依赖于细胞骨架及其结合蛋白组成的动力系统。
- 信息传递与代谢调节:细胞骨架参与细胞间的信息交流,并为多种酶提供附着位点,使代谢反应高效有序进行。
- 细胞分裂与分化:它是细胞分裂过程中必不可少的机械装置,同时在细胞分化过程中引导细胞形态的改变。
细胞骨架在不同生物中的特殊性:
- 动物细胞:由于缺乏细胞壁,动物细胞尤其依赖细胞骨架来维持形态及承受外力。
- 植物细胞:尽管有细胞壁支撑,细胞骨架仍不可或缺。它不仅负责胞质流动,还负责引导纤维素微纤丝的沉积方向,从而控制植物的生长方向。
- 原核细胞:虽然早期认为原核生物没有细胞骨架,但现代研究发现细菌也拥有与真核生物骨架蛋白同源的成分,参与细胞分裂和极性决定。
细胞骨架的异常与多种疾病相关。例如,硒缺乏会导致线粒体膜不稳定,影晌心肌细胞中骨架的稳定性;二氧化硅粉尘(矽肺)会通过破坏肺部吞噬细胞的溶酶体及相关的骨架系统导致细胞死亡。此外,抗肿瘤药物如秋水仙素通过干扰微管的动态平衡来抑制癌细胞的分裂。综上所述,细胞骨架是一个精巧而复杂的生命支撑系统,它使细胞成为一个既能保持相对稳定,又能根据环境需求迅速做出反应的有机统一整体。
【浙科版】细胞壁是一些细胞的最外层结构。植物的细胞壁主要由纤维素组成,木材、纸张、棉、麻的主要成分都是植物细胞壁中的纤维素。此外,植物细胞壁还含有果胶等多种物质。真菌和大多数原核细胞同样具有细胞壁,但组成物质和结构与植物不同。动物细胞没有细胞壁。细胞壁具有全透性,水、离子和其他分子都极易通过细胞壁的空隙,因此细胞壁与细胞的选择透过性无关。细胞壁的重要作用是保护细胞,维持细胞形态,加强细胞的机械强度。参天大树之所以能挺立,主要是靠死细胞的细胞壁支撑。此外,细胞壁也参与细胞间的相互粘连,是激素等化学信号传递的介质和通路。
细胞壁(Cell wall)是位于细胞膜外层的一种坚硬、刚性的结构,它广泛存在于植物、真菌、大多数原核生物(如细菌)及某些原生生物中,但在动物细胞中缺失。细胞壁不仅为细胞提供机械支持和形状,还起着保护细胞免受环境压力和渗透裂解的作用。不同生物类群的细胞壁化学成分差异显著,这是区分它们的重要依据:
- 植物细胞壁:主要由纤维素(Cellulose)组成,这是一种由葡萄糖单体构成的多糖。此外还含有果胶(Pectin)、**半纤维素(Hemicellulose)以及少量的蛋白质。在某些特化组织(如木质部)中,还会沉积木质素(Lignin)以增加硬度,或沉积木栓质(Suberin)**以起到防水作用。
- 细菌细胞壁:绝大多数细菌的细胞壁主要成分是肽聚糖(Peptidoglycan)。根据革兰氏染色反应,细菌可分为革兰氏阳性菌(具有厚肽聚糖层并含磷壁酸)和革兰氏阴性菌(具有薄肽聚糖层及含脂多糖的外膜)。
- 真菌细胞壁:主要成分是几丁质(Chitin)和葡聚糖(Glucans)。几丁质也是昆虫等节肢动物外骨骼的主要成分。
- 古细菌细胞壁:不含肽聚糖,可能含有假肽聚糖、多糖、糖蛋白或纯蛋白质。
- 藻类细胞壁:成分多样,包括纤维素、琼脂、褐藻胶或二氧化硅(如硅藻)。
- 特殊例外:支原体(Mycoplasma)是目前已知最小、最简单的原核生物,其显著特征是没有细胞壁。
植物细胞壁并非单一结构,而是随细胞生长分层形成的:
- 胞间层(Middle lamella):位于相邻两个细胞的最外层,主要成分是果胶,起着将相邻细胞“胶合”在一起的作用。
- 初生壁(Primary wall):在细胞生长过程中形成,位于胞间层内侧,较薄且富有弹性(含纤维素、半纤维素和果胶),允许细胞伸长。
- 次生壁(Secondary wall):细胞停止生长后,在初生壁内侧沉积形成。它比初生壁厚且坚硬,常含有木质素,起支持和保护作用。
细胞壁的核心功能:
- 支持与定型:细胞壁作为坚硬的外壳,维持细胞的特定形状,并支撑整株植物抵抗重力,挺立于空中。
- 保护作用:保护细胞免受机械损伤和病原体(如病毒、细菌)的侵染。
- 调节渗透压与防止裂解:当细胞处于低渗环境(如清水)中时,细胞会渗透吸水。细胞壁产生的胞壁压能阻止细胞无限膨胀,从而防止细胞因渗透作用而吸水涨破(裂解)。
- 信息交流与运输:植物细胞壁上具有纹孔(Pits),**胞间连丝(Plasmodesma)**可穿过纹孔连接相邻细胞的细胞质,成为细胞间物质运输和信息交流的重要通道。
- 全透性(Full permeability):与具有选择透过性的细胞膜不同,细胞壁是全透性的。水、离子和小分子物质可以自由通过细胞壁的空隙,因此它不是细胞这个系统的边界(系统的边界是细胞膜)。
合成机制:
- 在植物细胞分裂的末期,高尔基体产生的囊泡会在赤道板位置聚集并融合形成细胞板,最终发育为新的细胞壁。
- 纤维素的合成由嵌入细胞膜的纤维素合成酶复合物完成,该过程受到细胞质膜下微管的引导,决定了纤维素微纤丝的沉积方向。
实践应用与相关现象:
质壁分离(Plasmolysis):当活的成熟植物细胞置于高渗溶液(如 0.3g/mL 蔗糖溶液)中时,由于原生质层的伸缩性大于细胞壁的伸缩性,细胞会失水收缩,导致原生质层与细胞壁分离。
原生质体制备(酶解法):在植物细胞工程中,常用纤维素酶和果胶酶去除细胞壁,以获得裸露的、具有生活力的原生质体。
抗生素作用原理:青霉素等抗生素通过抑制细菌细胞壁(肽聚糖)的合成来达到杀菌效果。由于人及动物细胞没有细胞壁和肽聚糖,因此这类抗生素对人体细胞几乎没有毒性,但对没有细胞壁的支原体也无效。
质体(Plastid)是真核绿色植物和藻类细胞特有的细胞器,是细胞进行光合作用、合成及储存碳水化合物、脂质和蛋白质的场所。质体由双层膜包被,并具有一定程度的遗传独立性。
质体的起源与发育:所有质体都起源于前质体。前质体主要存在于植物的根尖、茎尖、胚及卵等幼龄细胞中,呈无色或淡绿色的球状体。随着细胞的生长和分化,前质体在不同环境条件下(如光照)可发育或转变成叶绿体、白色体或有色体。这些不同类型的质体在特定条件下也可以相互转化。
质体的主要类型:根据所含色素和生理功能的不同,质体主要分为三类。
叶绿体(Chloroplast)——“养料制造车间”
- 功能:绿色植物进行光合作用的场所。它能捕捉太阳能,将二氧化碳和水还原为富含能量的糖(如淀粉、蔗糖),同时释放氧气。
- 结构:
- 被膜:由外膜和内膜两层单位膜组成。
- 类囊体(Thylakoid):悬浮在基质中的扁平膜囊,分布着光合色素(叶绿素、类胡萝卜素)和光反应所需的酶。
- 基粒(Granum):许多圆盘状的类囊体有序叠摞而成,极大地扩展了膜面积。
- 基质(Stroma):内膜包围的液态物质,含有暗反应(碳反应)所需的酶。
- 分布特性:叶绿体在细胞中的分布随光照而变化。弱光下,其正面朝向光源以最大化吸收光能;强光下,侧面朝向光源以避免灼伤。
有色体(Chromoplast / Chromoplast)
- 成分:含有类胡萝卜素,包括黄色的叶黄素和红色的胡萝卜素。
- 功能:使花瓣、成熟果实、秋天落叶以及某些根(如胡萝卜)呈现出鲜艳的红、橙、黄色。这些颜色有助于吸引动物传粉或传播种子。
白色体(Leucoplast / Leucoplast)
- 特性:不含色素,呈无色,结构简单。
- 分布:多分布在植物不见光的部位,如根、茎的地下储藏器官。
- 分类(按储藏物质):
- 造粉体(Amyloplast):专门储藏淀粉,如马铃薯块茎中富含造粉体。
- 造油体(Elaioplast):专门储藏脂质(油脂)。
- 造蛋白体(Proteinoplast / Proteoplast):专门储藏蛋白质。
质体的半自主遗传特性:质体(尤其是叶绿体)被认为是半自主性细胞器,其依据如下。
- 遗传物质:拥有独立的环状 DNA 分子(质体 DNA),能自主复制。
- 合成系统:拥有自己的核糖体(70S 型)和 RNA,能合成自身所需的一部分蛋白质。
- 繁殖方式:不随细胞核分裂,而是通过二分裂(类似细菌)进行增殖。
内共生起源学说(Endosymbiont Theory):目前广泛接受的学说认为,质体起源于古代原始真核细胞吞噬的蓝细菌(蓝藻)。
- 质体 DNA 与蓝细菌 DNA 的序列高度相似。
- 其 70S 核糖体在结构上与细菌核糖体更为接近。
- 双层膜结构中,内膜与细菌质膜成分相似,外膜则与宿主细胞膜成分相近。
核心考点总结:
- 特有性:质体是植物细胞特有的,动物细胞完全没有。
- 含有色素的细胞器:质体(叶绿体、有色体)和液泡。
- 含有 DNA/RNA 的细胞器:质体(叶绿体)和线粒体。
- 功能统一:不同类型的质体通过分工合作(如叶绿体提供养料,白色体储存养料),共同维持植物的生长发育。
内质网(Endoplasmic Reticulum, ER)是真核细胞中面积最大、联系最广的膜性管道系统,由单层膜围成的扁平囊状或管状结构相互沟通而成。它在细胞内占据中心地位,通过直接连接、囊泡运输或细胞骨架锚定等方式与多种细胞器及结构紧密关联,共同构建了一个高效有序的代谢网络。
以下是吸附、紧靠或与内质网有直接关联的主要细胞器及结构,以及它们之间的相互作用:
细胞核(核膜):内质网膜与核膜的外膜直接相连。从进化角度看,核膜被认为是围绕核物质的特化内质网部分。两层核膜之间的腔隙(核周隙)与内质网腔也是相通的。
- 信息传递与协调:这种直接的结构连续性有利于遗传信息从细胞核(转录产生 mRNA)快速传递到细胞质中的内质网进行翻译和加工。
- 膜成分更新:细胞分裂末期,核膜的重建往往以与之相连的内质网膜为基础。
核糖体:**粗面内质网(RER)**的外表面吸附有大量的核糖体颗粒。
- 蛋白质合成与转运:核糖体是“生产蛋白质的机器”。分泌蛋白及多种膜蛋白在核糖体上起始合成后,通过信号肽引导转移至内质网,边合成边进入内质网腔内或定位在膜上。
- 初步加工:进入内质网的多肽链在此进行折叠、组装及糖基化修饰,形成具有一定空间结构的蛋白质。
高尔基体:内质网与高尔基体通过囊泡进行间接但频繁的联系。这种动态的物质转移被称为“膜流”。
- 分泌蛋白的运输通路:在分泌蛋白的合成过程中,内质网将初步加工的蛋白质包裹在囊泡中运输至高尔基体。
- 进一步加工与分拣:高尔基体对来自内质网的蛋白质进行分拣、分类、包装及进一步的化学修饰(如复杂的糖基化),随后再将其发送至细胞膜或溶酶体。
- 逆向运输(回收机制):若内质网驻留蛋白错误进入高尔基体,高尔基体可通过特定囊泡(如 COP I 包被小泡)将其运回内质网。
线粒体:在某些细胞中,内质网膜与线粒体外膜直接相连。这种特化的接触位点被称为内质网 - 线粒体连接。
- 能量供应:线粒体作为“动力车间”,为内质网上的蛋白质合成、加工及囊泡运输提供所需的 ATP。
- 脂质交换:内质网是合成磷脂的主要场所,可通过直接接触或磷脂交换蛋白(PEP)将脂质转运至线粒体膜上。
- 钙离子调节:内质网是细胞内最大的“钙库”,它与线粒体间的钙离子交换对细胞信号转导和能量代谢应答至关重要。
细胞膜(质膜):内质网膜向外直接连接细胞膜。此外,两者通过囊泡运输保持功能上的高度统一。
- 物质交换:内质网合成的膜蛋白和脂质最终通过囊泡融合的方式融入细胞膜,实现膜成分的更新。
- 信号传递:内质网作为细胞内庞大的膜性网络,通过与细胞膜的连接参与胞内外的信号沟通。
溶酶体与液泡:溶酶体(动物)和液泡(植物)中含有的多种水解酶最初是在内质网上的核糖体中合成的,并经过内质网的初步加工。
- 形成与补充:内质网特化或分离出的囊泡参与了液泡、溶酶体以及其他微体的形成。
- 细胞自噬:当内质网受损或发生应激时,受损的内质网碎片会被包裹形成自噬小体,最后与溶酶体(或液泡)融合并被降解。
细胞骨架:内质网被锚定并支撑在细胞骨架(主要是微管和微丝)上。
- 空间定位与形态维持:细胞骨架决定了内质网在细胞内的分布格局,例如通过马达蛋白沿着微管将内质网拉向细胞边缘。
- 物质运输轨道:从内质网出发前往高尔基体的囊泡通常由细胞骨架提供运输轨道。
胞间连丝(植物细胞特有):在高等植物细胞间,内质网可以延伸穿过胞间连丝,形成连接相邻细胞的连丝微管(Desmotubule)。
- 跨细胞运输与通讯:这一结构使得相邻细胞的内质网、细胞质及质膜交融在一起,成为植物细胞间物质运输(如代谢物、小分子蛋白、RNA)和信息传递的重要通道。
综上所述,内质网不仅是蛋白质和脂质的**“合成与加工车间”,更是细胞内膜系统、能量代谢系统及骨架支撑系统的枢纽**,通过与上述细胞器的紧密协作,保证了细胞生命活动的高效、有序进行。
观察叶绿体和胞质环流实验
这个实验是高中生物学中观察“活”细胞动态生命现象的核心实验。它通过观察绿色的叶绿体作为参照标影,来揭示肉眼看不见的胞质环流。
实验材料的深度选择:
- 黑藻 (Elodea):是观察胞质环流的首选材料。它的叶片极薄(仅两层细胞),叶肉细胞大且叶绿体清晰,非常适合直接制片观察。
- 藓类小叶:同样是理想材料,因为许多藓类的叶片仅由一层细胞组成,不需要做切片处理即可清晰观察。
- 菠菜叶:若使用菠菜,必须撕取下表皮并稍带些叶肉。这是因为表皮细胞通常不含叶绿体,只有叶肉细胞才含有大量叶绿体供观察。
关键的“预处理”:唤醒细胞。胞质环流的速度受环境影响显著。在实验前,通常需要将材料放在光照充足、**温度适宜(约 25℃)**的环境中培养 15-30 分钟。
- 光照:提供能量,激发代谢。
- 温度:提高酶活性,增加细胞质的流动性。
- 物理损伤:有时轻轻切伤部分叶片,也能刺激邻近细胞加快环流速度。
操作细节与注意事项:
- 保持有水:制作临时装片时,水分要充足。实验过程中材料绝不能干涸,否则细胞失水会影响代谢甚至导致死亡,使环流停止。
- 高倍镜观察:必须先在低倍镜下找到清晰的叶肉细胞,将其移至视野中央,再转换高倍镜并仅调节细准焦螺旋。
实验现象:叶绿体的“策略”移动。在显微镜下,你会发现叶绿体并非静止,而是沿着细胞壁随细胞质进行环形流动。
- 避光反应:这是一项精妙的适应。在弱光下,叶绿体会以椭球体的正面朝向光源以吸收更多光能;在强光下,它会转动方向,以侧面朝向光源,避免被高能量灼伤。
- 环流方向:在视野中观察到的环流方向(如顺时针)与装片中实际的流动方向是一致的。
讨论:结构的动态意义。
- 形态与功能:叶绿体呈扁平的椭球形,有利于通过改变位置和方向来调节受光面积。
- 流动的价值:胞质环流不仅能使叶绿体移动到最佳光照位置,还能加速细胞内营养物质的分配和代谢废物的运输,确保细胞内各项生化反应高效有序进行。
这个实验让你对“细胞是生命活动的基本单位”有了直观的动态理解。
细胞的生物膜系统
细胞的生物膜系统是一个由细胞膜、核膜以及内质网、高尔基体、线粒体、叶绿体、溶酶体等具膜细胞器共同构成的复杂网络。这些膜结构在化学组成和空间结构上高度相似,主要由磷脂双分子层构成基本支架,并镶嵌、贯穿有功能各异的蛋白质。
该系统的核心价值在于其精细的功能分工与高度的协调性:
- 屏障与稳定:细胞膜作为系统的边界,不仅保障了细胞内部环境的相对稳定,还通过选择透过性控制物质进出,并参与细胞间的信息交流。
- 区室化(功能化区域):生物膜将细胞质分隔成多个相对独立的区室,使得如葡萄糖合成与分解等互不相容的化学反应能同时高效进行,避免相互干扰。
- 代谢支持:膜结构提供了极其广阔的表面积,为多种酶提供了附着位点,从而支持复杂的生化反应高效有序地开展。
这些组分在结构和功能上是紧密联系的动态整体。例如,内质网膜可以与核膜、细胞膜直接相连;而在分泌蛋白的合成运输中,内质网、高尔基体和细胞膜通过囊泡的“出芽”与融合实现间接联系和膜成分的更新。
【浙科版】真核细胞内存在着丰富的膜结构,它们将细胞内部划分成相对独立的区室,保证多种生命活动高效、有序地进行。这些膜结构又是可以相互转化的,在结构和功能上构成一个统一整体。细胞膜和包括核膜在内的多种细胞器膜共同构成细胞的生物膜系统。生物膜中,内质网的面积最大,占了整个细胞生物膜的一半左右。内质网是细胞内除核酸以外一系列重要的生物大分子如蛋白质、脂质和糖类的加工或合成基地。这些生物大分子被合成后可在内质网完整、封闭的管道中快速运输。由于内质网膜向内与核膜连接,向外与细胞膜连接,使细胞核与细胞质及细胞膜的联系更为紧密,大大提高了物质交换的效率。由于生物膜具有一定的流动性,所以各组分间可以通过囊泡相互转化。内质网以出芽的形式形成囊泡,囊泡移动到高尔基体并与其融合。高尔基体同样以囊泡的形式与细胞膜融合,成为细胞膜的一部分。细胞膜也可以内陷形成囊泡回到细胞质中。囊泡的移动为多种细胞器的膜质成分提供物质来源,从而实现膜成分的更新。
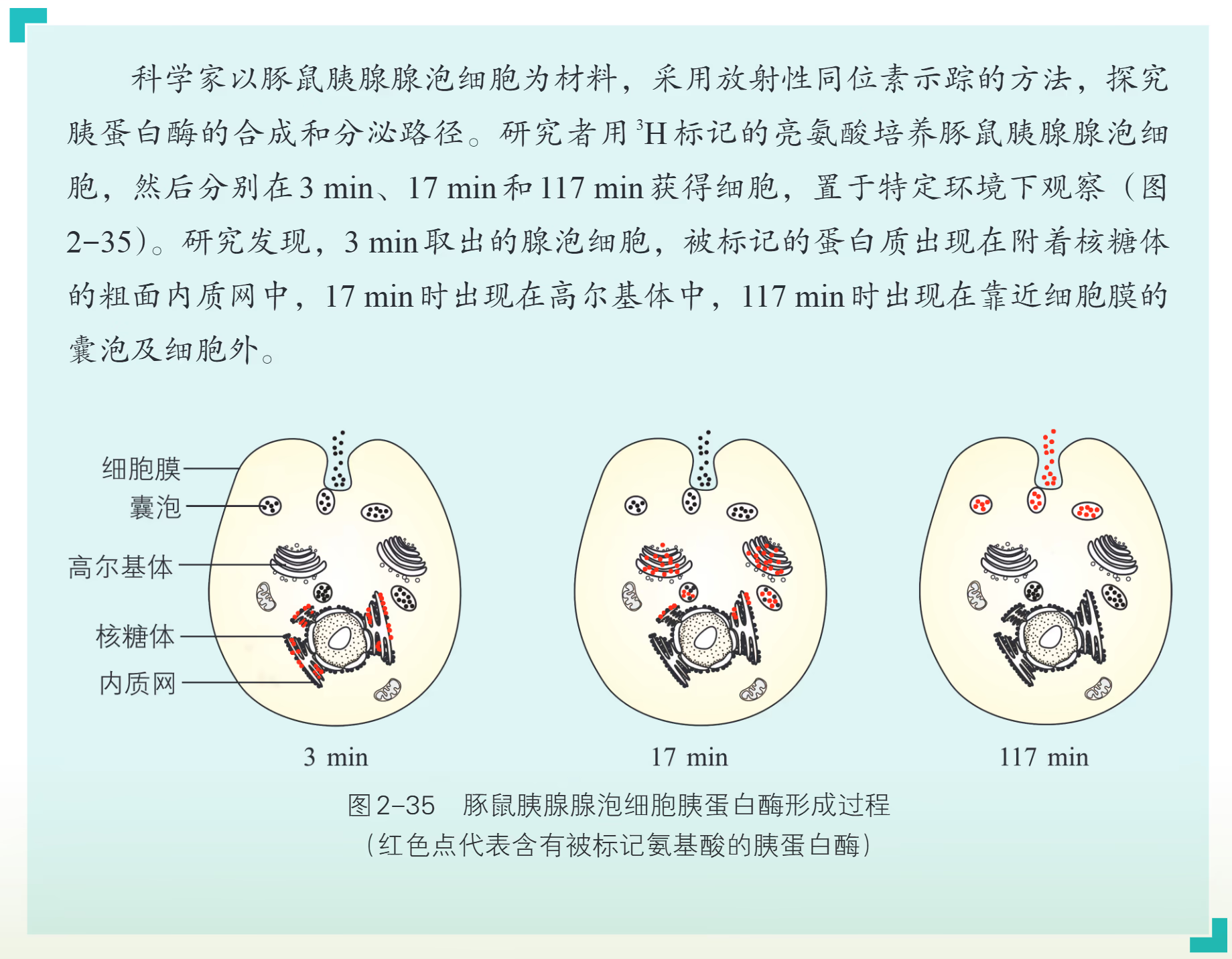
机体中有一类在细胞内产生,分泌到胞外发挥作用的蛋白质,称为分泌蛋白,如抗体、消化酶和一部分激素。分泌蛋白是如何产生、分泌的?
由上述小资料可知,胰蛋白酶的合成和分泌是在多种细胞结构的参与下进行的。首先,氨基酸在核糖体(附着于内质网上)中形成多肽,之后通过内质网的加工和运输,随囊泡转移至高尔基体。随后,高尔基体形成的囊泡包裹着蛋白质向细胞膜移动,将蛋白质分泌至胞外。在光面内质网合成的磷脂和胆固醇同样需要与高尔基体的联系,通过囊泡运入其他细胞器。
许多生物大分子在内质网和高尔基体等细胞器内合成或加工后,不能直接穿过生物膜,而是被包裹在囊泡里,并被运至特定部位。通过囊泡运输的物质主要有两类:一类是囊泡膜上的膜蛋白和脂质等,参与细胞器的组成并完成特定的细胞功能;另一类是囊泡包裹的内含物,如一些激素、酶等,这些物质有的被分泌到细胞外,有的参与细胞内大分子物质的降解等。一般来说,囊泡运输包括囊泡形成、运输和与特定部位膜的融合,其中囊泡与特定部位膜的融合是囊泡定向运输的关键。整个过程非常复杂,需要多种信号分子和细胞骨架的参与。
细胞在代谢过程中产生的残渣、功能异常的大分子以及衰老的细胞器等,被内质网或其他膜结构形成的囊泡包裹着,与溶酶体融合,进而被消化分解,产生的小分子物质有的被细胞重新利用,有的被排出细胞。
生物膜系统大大提高了细胞内物质运输的效率,加强了各组分间的交流,同时由生物膜形成的各区室使细胞具有相对独立的空间,保证了细胞各项生命活动高效、有序地进行。细胞内广阔的膜面积为多种酶提供了附着位点,为多种化学反应顺利进行提供了有利条件。所以,生物膜系统使细胞在结构和功能上成为一个统一的整体。
**囊泡(Vesicle)**是真核细胞中极其重要的动态膜性结构,被形象地比喻为细胞内的“深海潜艇”或“搬运工”。它们承担着细胞内大分子物质的定向运输、分选及分泌等核心任务。
基本结构:囊泡是由脂质双分子层(主要为磷脂)包围而成的封闭球形或椭球形小泡。由于其结构并不固定(始终处于产生与融合的动态中),因此通常不被视为一种相对稳定的“细胞器”。
化学成分:
- 膜成分:主要由磷脂和蛋白质构成,还含有糖类。在动物细胞中,胆固醇常存在于囊泡膜上,用于调节膜的流动性和增加稳定性。
- 内含物:囊泡内包裹着待运输的“货物”,如蛋白质(激素、酶、抗体)、脂质、核酸或代谢废物等。
结构特性:囊泡具有一定的流动性,这是其能够与细胞膜及其他细胞器膜发生融合的结构基础。
根据其外被蛋白(Coat protein)的不同,目前了解较多的有三类有被囊泡:
- COP II 有被囊泡:产生于粗面内质网,负责介导从内质网到高尔基体的“顺向转运”。
- COP I 有被囊泡:产生于高尔基体,负责将内质网驻留蛋白等“逃逸”物质回收转运回内质网(逆向转运),也参与高尔基体内部的物质运输。
- 网格蛋白有被囊泡(Clathrin-coated vesicle):可产生于高尔基体或细胞膜。它们介导从高尔基体向溶酶体或质膜的运输,以及受体介导的胞吞作用。
- 特殊类型:包括突触小泡(存储神经递质)、分泌囊泡(储存分泌蛋白)以及参与细胞自噬的自噬体等。
囊泡的运行是一个精密调控的过程,主要分为以下四个阶段:
- 出芽(Budding):在特定部位的膜(供体膜)受信号激发,包被蛋白组装并促使膜向外凸起或向内凹陷。**发动蛋白(Dynamin)**在囊泡颈部聚合形成环状,通过水解 GTP 产生的能量将囊泡从母体膜上“掐断”。
- 运输(Transport):囊泡离开供体膜后,通常依赖细胞骨架(主要是微管)作为轨道。马达蛋白(如驱动蛋白 Kinesin 或动力蛋白 Dynein)利用 ATP 提供的能量,牵引囊泡沿轨道定向移动。
- 锚定与系留(Docking & Tethering):囊泡表面特有的 Rab 蛋白被靶膜上的束缚蛋白识别并结合,使囊泡停靠在特定的目的地。
- 融合(Fusion):涉及 SNARE 蛋白复合体的相互作用。囊泡上的 v-SNARE 与靶膜上的 t-SNARE 像绞车一样互相缠绕,克服膜表面的静电斥力,使两层膜紧密接触并最终融合,将其中的“货物”释放。
核心生理功能:
蛋白质的分泌途径:这是囊泡最重要的功能之一。分泌蛋白在核糖体合成后,依次经过内质网(加工、折叠)、高尔基体(再加工、分类、包装),全程通过囊泡进行转运,最后通过胞吐释放到细胞外。
胞吞与胞吐作用:
- 胞吞(内吞):细胞通过囊泡摄取大分子或颗粒物(如白细胞吞噬病菌),胞吞形成的囊泡通常会与溶酶体融合并被降解。
- 胞吐(外排):细胞通过囊泡排出分泌物或代谢废物(如神经递质的释放)。
膜成分的更新与转化:囊泡在不同膜性细胞器之间穿梭,实现了生物膜系统在成分和结构上的相互转化与代谢更新。
细胞间信息交流:某些细胞能分泌包含蛋白质、核酸的外泌体(一种细胞外囊泡),通过血液运输到其他部位,介导远距离的信息传递。
囊泡转运的精准性对维持细胞稳态至关重要,转运障碍可能导致多种疾病:
- 糖尿病:胰岛素合成后若囊泡运输出现故障,或无法准确与细胞膜融合释放,会导致血糖控制失调。
- 矽肺:肺部吸入矽尘后,溶酶体囊泡膜受损导致水解酶泄露,破坏细胞结构,最终导致肺功能受损。
- 神经系统疾病:突触小泡释放神经递质的异常与多种神经系统障碍有关。
- 病毒感染:某些病毒(如流感病毒、HIV)会利用受体介导的胞吞作用,“劫持”细胞囊泡进入细胞内部。
总结来说,囊泡不仅是物质运输的载体,更是维持真核细胞内部高度有序化、区域化生理活动的关键纽带。
在实际应用中,人工合成的透析膜正是模拟了生物膜的选择透过性,能够像“人工肾”一样过滤血液中的代谢废物,同时保留蛋白质等大分子,从而救治肾功能障碍患者。生物膜系统在细胞的生命活动中作用极为重要(课本原话):
- 第一,细胞膜不仅使细胞具有一个相对稳定的内部环境,同时在细胞与外部环境进行物质运输、能量转化和信息传递的过程中起着决定性的作用。
- 第二,许多重要的化学反应需要酶的参与,广阔的膜面积为多种酶提供了附着位点。
- 第三,细胞内的生物膜把各种细胞器分隔开,如同一个个小的区室,这样使得细胞内能够同时进行多种化学反应,而不会互相干扰,保证了细胞生命活动高效、有序地进行。
人工合成的膜材料已用于疾病的治疗。例如,当肾功能发生障碍时,由于代谢废物不能排出,病人会出现水肿、尿毒症。目前常用的治疗方法,是采用透析型人工肾替代病变的肾行使功能,其中起关键作用的血液透析膜就是一种人工合成的膜材料。当病人的血液流经人工肾时,血液透析膜就能把病人血液中的代谢废物透析掉,让干净的血液返回病人体内。
细胞核的结构与功能
细胞核是真核细胞的核心细胞器,被誉为细胞的“大脑”或“控制中心”,其精巧的结构直接决定了其强大的调控功能。以下是根据来源对细胞核结构与功能的详细展开:
核膜(核被膜):双层屏障与连续性
- 结构:核膜由两层同心的磷脂双分子层组成(内膜和外膜),将核内物质与细胞质严格分开。
- 连续性:核外膜通常与粗面内质网相连,其表面也常附着有核糖体。
- 核纤层:内核膜内表面有一层由蛋白质纤维组成的网状结构,称为核纤层,起支撑核膜和固定染色质的作用。
核孔:严格的“海关”通道
- 功能:实现核质之间频繁的物质交换和信息交流。
- 选择性:它并非简单的物理孔洞,而是由约 30 种蛋白质构成的核孔复合体。它允许水、离子等小分子自由通过,但对蛋白质、RNA 等大分子进出有严格的选择性。
- 代谢关联:代谢越旺盛、蛋白质合成越频繁的细胞,核孔的数目通常越多。
核仁:核糖体的“组装车间”
- 功能:是 rRNA(核糖体 RNA)合成、加工以及核糖体亚单位装配的场所。
- 动态性:核仁没有膜包被,在细胞分裂过程中会周期性地消失和重建。
染色质与染色体:遗传物质的两种形态
- 成分:主要由DNA 和蛋白质(主要是组蛋白)组成。
- 包装:DNA 缠绕在组蛋白上形成核小体(最基本的包装单位),再进一步折叠形成染色质丝。
- 转变:在间期,遗传物质呈细丝状的染色质形态,有利于 DNA 的复制和转录;在分裂期,它高度螺旋化、缩短变粗成为染色体,有利于遗传物质的平均分配。
细胞核的核心功能:
- 遗传信息库:细胞核是绝大多数遗传物质(DNA)储存和复制的场所。它包含了生物体发育和性状表达的全部“蓝图”。
- 代谢与遗传的控制中心:
- 控制性状:如美西螈核移植实验和伞藻实验证明,生物体的性状(如肤色、伞帽形状)是由细胞核决定的。
- 控制代谢:变形虫切割实验显示,失去细胞核的细胞部分无法摄食和生长,最终死亡,证明细胞核维持着正常的生命活动。
- 指导合成:细胞核通过转录产生 mRNA,mRNA 穿过核孔进入细胞质,指导核糖体合成蛋白质,从而调控细胞的所有生理活动。
值得注意的是,并非所有真核细胞都终身拥有细胞核。例如,哺乳动物成熟的红细胞和高等植物成熟的筛管细胞为了适应运输功能,在成熟过程中会失去细胞核。
这两个经典实验是生物学中探究细胞核功能的核心证据,通过对单细胞生物的操作,证明了细胞核是细胞代谢和遗传的控制中心。以下是根据来源进行的详细展开讲解:
变形虫(Amoeba)切割实验:变形虫是一种结构简单的单细胞原生动物,具有伪足,能通过胞吞摄食,是研究细胞核与细胞质关系的理想材料。
- 切割操作:科学家用纤细的玻璃针将一个变形虫切割成两半,一半含有细胞核(有核部分),另一半不含细胞核(无核部分)。
- 观察结果:
- 有核部分:能正常生活,照样摄食,对刺激仍有反应,且失去的伸缩泡可以再生,还能进行生长和分裂。
- 无核部分:虽然能短期生存(消化已吞噬的食物),但不能摄取新食物,对外界刺激不再发生反应,且电镜下可见高尔基体、内质网等细胞器退化,最终在一段时间后(约 20 天内)死亡。
- 核植入实验(自身前后对照):如果在去核后的三天内(或 10 分钟内),向无核部分植入一个完整的新细胞核,变形虫的生命活动(如摄食、生长、分裂、应激性等)又会陆续恢复,重新开始正常生活。
- 结论:变形虫的分裂、生长、再生及对刺激的反应等生命活动均受细胞核控制。
- 深度洞察:
- 该实验还说明了细胞只有保持结构的完整性,才能完成正常的生命活动。
- 去核后仍能短期生存的原因:是因为细胞质中已合成的蛋白质和由细胞核转录出的 mRNA 仍可继续发挥作用一段时间。
- 实验评价:该实验既有相互对照(有核与无核对照),又有自身前后对照(去核后重新植入核)。
伞藻(Acetabularia)实验:伞藻是生活在热带海域的大型单细胞绿藻(长 2~5cm),由伞帽、伞柄和假根三部分构成,细胞核位于基部的假根内。实验常选用“伞形帽”和“菊花形帽”两种伞藻作为材料。
伞藻“嫁接”实验:
- 过程:科学家切除两种伞藻的帽,并将它们的伞柄进行互换“嫁接”。即:将伞形伞藻的柄嫁接到菊花形伞藻的假根上,反之亦然。
- 现象:一段时间后,嫁接体长出的新伞帽形状与提供假根的伞藻一致,而与提供伞柄的伞藻无关。
- 进阶细节:有时第一次再生的伞帽呈中间类型(不完全形),但将其切除后第二次再生的伞帽则完全符合假根的类型。这是因为原伞柄中残留了由原细胞核转录出来的遗传信息(mRNA)。
- 初步结论:伞藻“帽”的形状是由假根决定的。
伞藻核移植实验
- 目的:为了排除假根中除细胞核以外的其他物质(如细胞质成分)对实验的影响,科学家进一步进行了核移植实验。
- 过程:将菊花形伞藻的细胞核取出,移植到去核并去帽的伞形伞藻的假根中。
- 现象:最终得到的再生伞藻长出了与供核亲本一致的菊花形帽。
- 结论:伞藻“帽”的形状受细胞核控制。
总结与核心考点:
- 控制中心地位:这两项实验共同证明了细胞核是细胞代谢和遗传的控制中心。
- 遗传信息库:细胞核之所以能成为控制中心,是因为它含有遗传物质 DNA,DNA 储存着细胞进行生命活动的“蓝图”。
- 应用延伸(克隆技术):
- 美西螈核移植实验进一步证实了动物细胞核具有全能性(虽然体细胞克隆比胚胎细胞克隆难度大得多)。
- 著名的**克隆羊“多莉”和克隆猴“中中”“华华”**的诞生,本质上都是这些单细胞核移植实验在高等动物身上的大规模应用。
- 结构适应功能:在这些实验中,细胞表现出明显的极性(如涡虫再生由前向后呈梯度递减),这体现了生命系统内部精密的调控机制。
载体蛋白在核膜上分布是正确的。 在生物学中,核膜并不是一个简单的封闭屏障,而是一个高度复杂的结构。根据提供的信息,载体蛋白在核膜上的分布及其相关作用可以从以下几个维度来理解:
核孔复合体被视为“特殊的载体蛋白”:核膜上最重要的运输结构是核孔复合体(NPC)。
- 性质归类:来源明确指出,核孔复合体可看作是一类特殊的载体蛋白,它构成了细胞核与细胞溶胶之间物质交换的通道。它也被描述为一种特殊的跨膜运输蛋白复合体。
- 运输功能:核孔复合体具有双功能性,既可以介导入核转运,也可以介导核输出。对于蛋白质(如 DNA 聚合酶、组蛋白)和 RNA 等大分子的运输,它表现出类似于载体蛋白的选择性主动运输特征,需要消耗能量(通常由 GTP 水解提供)并依赖于特定的信号识别机制(如核定位信号 NLS)。
核膜(脂双层)本身分布有转运蛋白:除了核孔之外,核膜的磷脂双分子层结构中也分布着多种膜蛋白。
- 小分子运输:虽然核孔允许小分子自由扩散,但研究表明,一些小分子物质和无机盐离子等也可以通过核膜(而非仅仅核孔)进出细胞核,这意味着核膜上存在相应的离子通道或载体蛋白。
- 外核膜的特殊性:核外膜在形态和生化性质上与粗面内质网膜非常接近,并与之相连续。因此,内质网膜上分布的许多蛋白质(包括参与蛋白质加工、脂质合成或物质转运的载体蛋白)在核外膜上也有分布。
核内膜的特有蛋白:核内膜具有独特的蛋白质组成,例如核纤层蛋白受体。这些蛋白虽然不一定都行使“载体”功能,但它们是核内膜上重要的膜整合蛋白,负责将核纤层锚定在内膜上,维持核的结构完整性。
总结:在高中及大学生物学语境下,核孔复合体常被视为一种具备载体蛋白特征的复杂结构;同时,由于核膜与内质网的结构连续性,常规意义上的载体蛋白确实分布在核膜(尤其是外核膜)上。
在真核生物细胞中,核仁(nucleolus)是细胞核内最显著的结构,通常呈现为球形或椭球形的致密小体。以下是关于核仁结构与功能的详细展开:
- 无膜包被结构:核仁是一个没有膜包被的细胞器。在光学显微镜下,由于其折光性强且容易被碱性染料(如苏木精、洋红、碘液等)染成深色,因此在核内清晰可见。
- 分子组成:核仁主要由核糖体 RNA(rRNA)、蛋白质以及编码 rRNA 的 DNA 片段(rDNA)组成。
- 超微结构分区:在电子显微镜下观察,核仁可进一步区分为不同的功能区,主要包括:
- 纤维中心(致密纤维组分):由新转录的 rRNA 前体构成,是 RNA 合成的部位。
- 颗粒组分:由正在加工成熟的核糖体亚单位前体颗粒组成,是起始组装核糖体的部位。
- 核仁组织者区(NOR):核仁是由特定染色体上的次级缢痕处,即“核仁组织者区”汇集形成的,该区域含有多拷贝的 rRNA 基因。在人类细胞中,这些基因分布在 10 个集群中,位于 5 对不同染色体的末端附近。
- 高度动态性:核仁是一个周期性消失和重建的结构。在有丝分裂前期,随着染色质高度螺旋化形成染色体,核仁会逐渐解体消失;当分裂完成进入末期,子细胞中又会重新组装形成新的核仁。
核仁的主要功能:
- rRNA 的合成与加工:核仁是 rRNA(核糖体 RNA)转录、加工(如加帽、多腺苷酸化、剪接)和修饰的重要场所。
- 核糖体亚基的装配:核仁被形象地称为“产生核糖体的工厂”。在这里,rRNA 与从细胞质经核孔进入核内的核糖体蛋白结合,装配成核糖体的大、小亚基。
- 遗传信息的转运中介:组装好的核糖体亚基随后通过核膜上的核孔排入细胞质,并在细胞质中最终组装成功能完整的核糖体,用于指导蛋白质的合成。
- 调节核糖体数量:核仁还参与调控细胞中核糖体总量的变化,以适应细胞代谢的需求。
核仁的大小、数目和形态随生物种类、细胞类型及代谢状态的不同而有显著差异。
- 代谢旺盛的标志:在蛋白质合成旺盛、生长活跃的细胞中(如癌细胞、胚胎细胞、某些内分泌细胞),核仁通常体积较大且数目较多。
- 代谢微弱的情况:相反,在代谢活动较弱的细胞中,核仁则相对较小或不明显。
- 例外情况:哺乳动物成熟的红细胞和高等植物成熟的筛管细胞在发育过程中失去了细胞核,因此也不含有核仁。
综上所述,核仁作为细胞核内的“信息加工与机器装配车间”,通过控制核糖体的生物发生,在细胞的遗传表达和蛋白质代谢中发挥着中心调控作用。
在真核生物细胞中,核孔复合体(Nuclear Pore Complex, NPC)是嵌入在核被膜上的巨大蛋白质结构,它不仅仅是一个简单的孔洞,更是控制细胞核与细胞质之间物质交换的“精精密闸门”。
核孔复合体是跨越核被膜内、外两层膜的复杂结构,其结构极其庞大且精细。
- 膜融合位点:核孔位于核被膜内、外两层膜相互融合形成的环形开口处。
- 蛋白质组成(核孔蛋白):一个核孔复合体由约 30 种不同的蛋白质(统称为核孔蛋白,Nucleoporins)组成,每种蛋白质都有多个拷贝。
- 超微结构特征:
- 外形与尺寸:在电镜下观察,核孔复合体呈现八角形对称的结构,直径约为 90~120nm。
- 颗粒与纤维:核孔复合体两侧伸出蛋白质纤维。在细胞质侧,纤维向外延伸形成胞质丝;在核质侧,纤维汇聚形成类似捕鱼笼的核篮(Nuclear basket)结构。
- 中央运输通道:核孔中心充斥着由核孔蛋白构成的无序、柔软的蛋白质网状结构(被形象地比作“海带床”或凝胶网格)。
核孔控制着物质在核质与胞质间的双向交换,其转运方式分为被动扩散和主动转运。
- 被动扩散(非选择性):
- 核孔中央存在充满水的通道,允许水分子、离子以及直径小于 9nm 的小分子(如核苷酸、单糖等)自由进出。
- 主动转运(选择性):
- 大分子的关卡:对于蛋白质(如聚合酶、组蛋白)和核糖核酸(RNA、核糖体亚基)等大分子,核孔表现出高度的选择性,必须依靠特定的信号和受体才能通过。
- 入核转运:蛋白质需携带核定位信号(NLS)。这种信号通常由一串带正电荷的赖氨酸或精氨酸组成,被胞质中的**输入蛋白(Importin)**识别并引导至核孔。
- 出核转运:在核内合成的 mRNA、tRNA 和组装好的核糖体亚基需携带核输出信号(NES)。这些信号被**输出蛋白(Exportin)**识别,穿过核孔排入细胞质。
核孔的动态性与数量调节:
- 与代谢的关系:核孔的数目并非固定不变,而是随细胞代谢状态调整。代谢旺盛、蛋白质合成频繁的细胞(如浆细胞、卵母细胞),核孔数目显著增多。例如,典型的哺乳动物细胞核约有 3000~4000 个核孔,而两栖类卵母细胞可达百万个。
- 细胞分裂中的变化:核孔是一个动态结构。在有丝分裂前期,随着核膜解体,核孔复合体解聚并消失;在末期核膜重建时,核孔蛋白重新装配并恢复功能。
核孔作为细胞核的“门禁系统”,具有以下关键意义:
- 实现核质交流:保证遗传信息的转录产物(RNA)及时运出,以及代谢所需的蛋白质及时运入。
- 维持区域化:防止核质中的成熟分子与胞质中的分子发生不必要的混合,保证细胞生命活动的高效与有序。
综上所述,核孔复合体通过其独特的物理屏障(网状结构)和化学识别系统(受体与信号机制),精确调控着真核细胞遗传表达的空间分隔与时间序列。
细胞的衰老与死亡
细胞的衰老(Cell Aging / Senescence)是细胞生命进程中的自然规律,表现为细胞生理状态和化学反应发生复杂变化,最终导致形态、结构和功能的退行性变化。
衰老细胞的主要特征。衰老细胞在形态和生化层面具有显著特征:
- 形态变化:细胞内水分减少,导致细胞萎缩、体积变小,新陈代谢速率减慢。
- 结构变化:细胞核体积增大,核膜内折,染色质收缩、染色加深。细胞膜的通透性改变,物质运输功能降低。
- 生化变化:细胞内多种酶的活性降低(如黑色素合成相关的酪氨酸酶活性降低导致白发)。细胞内色素(如脂褐质)逐渐积累,妨碍物质交流。线粒体数量减少但体积增大,呼吸速率减慢。
细胞衰老的机制假说:
- 自由基学说:细胞代谢产生的异常活泼的带电分子(自由基)会攻击磷脂、DNA 和蛋白质。攻击磷脂会损伤生物膜并产生更多自由基;攻击 DNA 可能引起基因突变;攻击蛋白质使活性下降。
- 端粒学说:每条染色体两端都有端粒(DNA-蛋白质复合体),其随细胞分裂而逐次缩短。当端粒缩短到一定限度,端粒内侧的正常基因序列受到损伤,导致细胞活动异常并走向衰老。
细胞衰老与个体衰老的关系:
- 单细胞生物:细胞衰老即是个体衰老。
- 多细胞生物:个体衰老是组成个体的细胞普遍衰老的过程,但年轻机体内也有衰老细胞(如皮肤角质层),老年机体内也有幼嫩细胞(如造血干细胞)。
细胞的死亡(Cell Death)。细胞死亡主要包括细胞凋亡(程序性死亡)和细胞坏死。
细胞凋亡 (Apoptosis)
- 定义:由基因决定的细胞自动结束生命的过程,属于程序性死亡。
- 形态学过程:细胞皱缩、凝聚,核膜破裂,DNA 断裂成片段,最终形成由质膜包裹的凋亡小体。凋亡小体随后被邻近细胞或巨噬细胞吞噬消化,不引发炎症反应。
- 分子机制:涉及胱天蛋白酶 (Caspase) 家族的级联反应。受 Bcl-2 家族蛋白(如促凋亡的 Bax/Bak 和抗凋亡的 Bcl-2)的精密调控。
- 生物学意义:塑造组织结构(如胎儿手指分离、蝌蚪尾部消失)、清除衰老或受损细胞、维持内环境稳态。
细胞坏死 (Necrosis)
- 定义:在种种不利因素(如急性损伤、毒素)影响下引起的被动死亡。
- 特征:细胞肿胀、破裂,内含物外溢,常引发局部的炎症反应。
溶酶体的发生(Biogenesis of Lysosomes)。溶酶体是真核细胞中由单层膜包围的细胞器,被称为细胞的“消化车间”。
- 结构成分:含有 60 种以上的酸性水解酶,其内部 pH 值约为 5。
- 发生过程 (M6P 途径):
- 溶酶体酶在核糖体上合成后进入内质网进行初步加工。
- 运输至高尔基体,在此其糖链被磷酸化,形成特殊的标记——6-磷酸甘露糖 (M6P)。
- 高尔基体反面网络 (TGN) 上的 M6P 受体识别该信号,将酶分选并包装到转运囊泡中。
- 囊泡与晚期内体融合,最终发育成熟为具有活性的溶酶体。
细胞自噬(Autophagy)。细胞自噬是细胞降解自身受损组分并进行物质再循环的关键机制。
基本过程:
- 形成自噬体:由内质网等结构形成的隔离膜包绕待降解的细胞器(如衰老的线粒体)或蛋白质,形成双层膜结构的自噬体。
- 融合与降解:自噬体与溶酶体融合形成自噬溶酶体。
- 回收利用:溶酶体内的水解酶将内含物分解为氨基酸、核苷酸等小分子,释放到细胞质基质中被重新利用。
自噬的分类:
- 巨自噬 (Macroautophagy):最常见的形式,涉及完整自噬体的形成。
- 微自噬 (Microautophagy):溶酶体或液泡膜直接内陷包裹胞质物质。
- 分子伴侣介导的自噬 (CMA):具有特定序列的蛋白质被分子伴侣识别后直接通过溶酶体膜上的受体进入内部。
生理意义:
- 维持稳态:清除功能异常的蛋白质和损伤细胞器,防止代谢废物堆积。
- 应对逆境:在饥饿条件下通过降解自身物质产生能量和营养以维持生存。
- 免疫防御:降解侵入细胞的病毒和细菌。
在一定条件下,激烈的细胞自噬也可能导致程序性细胞死亡,即自噬性细胞死亡。
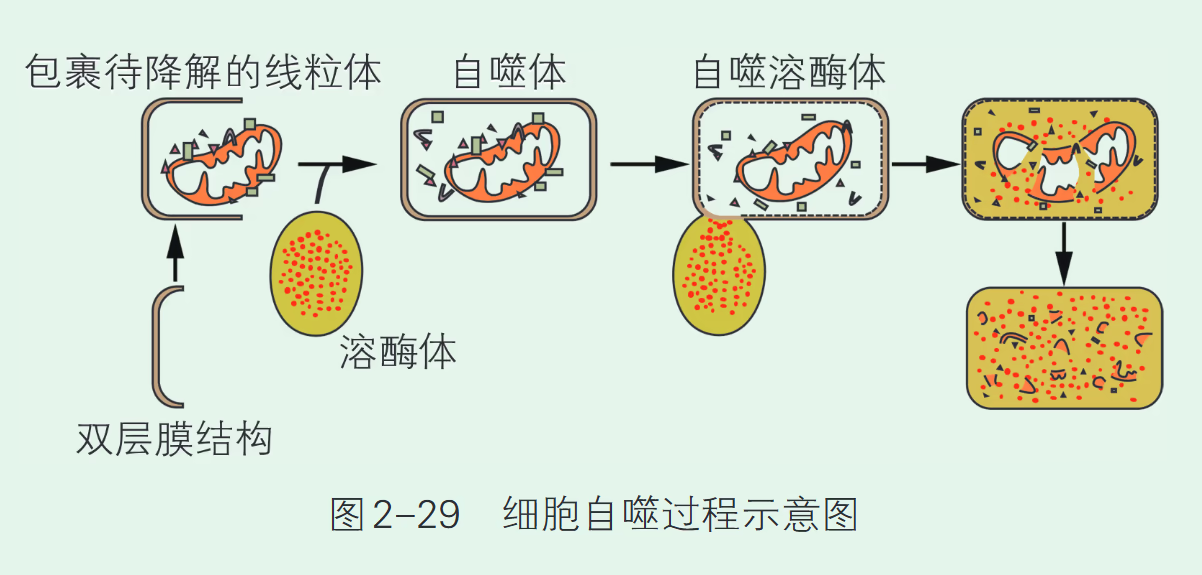
【浙科版】自噬(autophagy)是细胞在溶酶体的参与下降解细胞自身物质的过程。细胞中由内质网等结构形成双层膜的膜泡,并逐渐扩展,包裹待降解的细胞器或其他内含物,然后闭合形成自噬体,进而与溶酶体融合,形成自噬溶酶体,内含物被溶酶体内多种水解酶消化分解。自噬是细胞的自我保护机制。一方面,细胞在代谢过程中不可避免地产生一些代谢废物和功能异常的物质,如结构异常的蛋白质、衰老的线粒体等。这些物质的堆积会影响细胞正常的生命活动,所以需要通过细胞自噬及时清理。另一方面,细胞在营养物质缺乏、低氧等特殊环境下,通过自噬降解自身蛋白质或细胞器,为维持生存提供基本原材料和能量。此外,自噬还能使细胞对抗病毒和细菌等病原体的侵染。所以,自噬是细胞内的重要生理机制,对细胞抵抗逆境、对抗病原体和维持细胞内环境稳态等都具有重要的意义。
细胞的物质运输
水进出细胞的原理和质壁分离实验
根据你提供的教材内容和相关资料,我为你详细讲解水进出细胞的原理。
【沪科技版】取少量紫红色的高锰酸钾颗粒投放到盛有清水的玻璃杯中,会观察到紫红色的高锰酸钾慢慢地向周边散开。物质(高锰酸钾)以自身分子运动为动力,从高浓度区域向低浓度区域移动,这种移动方式称为扩散。
长颈漏斗实验解析:
- 液面升高的原因:单位时间内透过半透膜进入漏斗的水分子数量多于渗出的数量,这种溶剂分子的定向扩散即为渗透作用。液面不会无限升高,当管内水柱产生的静水压力与渗透压平衡时,水分进出达到动态平衡。
- 更换纱布的结果:纱布孔隙很大,不属于半透膜,蔗糖和水分子都能自由通过,因此液面不会升高。
- 两侧浓度相等的结果:没有浓度差(渗透压差),水分子进出平衡,液面保持不变。
水进出细胞的核心原理:渗透作用
- 定义与条件:水分子顺着相对含量梯度的扩散称为渗透。发生渗透必须具备两个条件:一是有半透膜,二是膜两侧存在浓度差。
- 分子基础:水分子极小,可以通过磷脂分子的间隙(自由扩散),但更多是借助膜上的水通道蛋白以协助扩散方式进出,这极大地提高了运输速率。
- 驱动力:水总是从高水势(低溶质浓度)流向低水势(高溶质浓度)。
哺乳动物红细胞的吸水与失水:
- 红细胞膜的性质:红细胞膜相当于一层半透膜,血红蛋白等大分子有机物无法滤过。
- 环境对形态的影响:
- 低渗环境:外界浓度小于细胞质浓度,细胞吸水膨胀甚至涨破(溶血)。由于红细胞没有细胞壁支撑,它极易受渗透压波动的影响。
- 等渗环境:外界浓度等于细胞质浓度,水分进出平衡,细胞维持正常形态。临床补液使用0.9% 的生理盐水正是为了维持这种平衡。
- 高渗环境:外界浓度大于细胞质浓度,细胞失水皱缩。
植物细胞的特殊性:
- 原生质层:由细胞膜、液泡膜及两层膜之间的细胞质组成,它相当于一层半透膜。
- 质壁分离:在高渗溶液中,原生质层因失水收缩,由于其伸缩性大于细胞壁,导致两者分离。
- 膨压作用:在低渗环境中,细胞吸水使液泡体积增大,产生向外的膨压(Turgor pressure),使植物挺拔;细胞壁则限制了细胞的过度膨胀,防止破裂。
植物细胞能发生质壁分离,关键在于其独特的结构——原生质层。原生质层由细胞膜、液泡膜以及两层膜之间的细胞质组成。在渗透作用中,原生质层整体相当于一层半透膜,而细胞壁则是全透性的。由于原生质层的伸缩性远大于细胞壁的伸缩性,当细胞失水时,两者就会发生分离。
实验的原理与条件:
- 渗透原理:当细胞液浓度小于外界溶液浓度时,细胞通过渗透作用失水,发生质壁分离;反之,当已分离的细胞置于低渗溶液中时,细胞吸水,发生质壁分离复原。
- 必备条件:
- 细胞必须是活的成熟植物细胞,且具有大液泡。
- 外界溶液必须与细胞液存在浓度差,且溶质分子不能轻易穿过原生质层(如高浓度蔗糖溶液)。
实验操作与观察指标:实验通常选用紫色洋葱鳞片叶外表皮细胞,因为其液泡含有色素,便于在显微镜下观察。
- 观察流程:首先在清水中观察正常状态;随后滴加 0.3g/mL 蔗糖溶液引发质壁分离;最后滴加清水观察复原。
- 变化特征:质壁分离时,液泡由大变小,紫色变深,原生质层与细胞壁逐渐分离。复原时,液泡由小变大,紫色变浅,原生质层恢复原状。
联系:在植物生物学中,原生质体(Protoplast)与原生质层(Protoplast layer)是两个极易混淆但本质不同的概念。从底层结构到应试应用,它们的区别可以总结为“整体与局部”、“遗传中心与功能膜”的差异。
原生质体(Protoplast):植物细胞的“活体整体”
- 概念:原生质体是指植物细胞除去细胞壁后剩余的全部生活物质。它是一个完整的生命单位,被视为一个“裸露的细胞”。
- 组成:包括细胞膜、细胞质和细胞核三部分。
- 存在条件:在自然状态下,它被细胞壁包围;在人工条件下,通过酶解法(纤维素酶和果胶酶)可以获得裸露的原生质体。
原生质层(Protoplast layer):成熟植物细胞的“选择透过性屏障”
- 概念:原生质层是成熟植物细胞(具有大液泡)中,起到半透膜作用的功能层。
- 组成:包括细胞膜、液泡膜以及两层膜之间的细胞质。
- 特别注意:原生质层不包括细胞核和液泡内的细胞液。
| 比较项目 | 原生质体 (Protoplast) | 原生质层 (Protoplast layer) |
|---|---|---|
| 所属关系 | 整体概念:细胞壁以内的全部结构 | 局部概念:成熟植物细胞的一部分 |
| 是否含细胞核 | 包含细胞核 | 不含细胞核 |
| 是否含细胞液 | 包含液泡及其中的细胞液 | 不含液泡内部的细胞液 |
| 核心功能 | 表现全能性,用于细胞工程和杂交 | 相当于半透膜,用于渗透吸水/失水 |
| 适用对象 | 所有的生活细胞 | 特指具有中央大液泡的成熟植物细胞 |
渗透作用与质壁分离(原生质层的舞台):在探究植物细胞吸水和失水的实验中,原生质层是核心。
- 半透膜特性:原生质层具有选择透过性,能让水分子通过而阻止蔗糖等溶质通过。
- 伸缩性差异:质壁分离的内在原因是原生质层的伸缩性大于细胞壁的伸缩性。当细胞失水时,原生质层由于具有高度伸缩性而收缩,导致其与刚性的细胞壁分离。
- 应试术语:质壁分离中的“质”指的就是原生质层,而非原生质体。
植物细胞工程(原生质体的舞台):在植物体细胞杂交技术中,原生质体是操作的对象。
- 去壁目的:由于细胞壁阻碍了细胞间的杂交/融合,必须先用酶解法去除细胞壁以获得原生质体。
- 特性:原生质体虽然没有细胞壁,但仍保持着细胞的全能性,在适宜条件下可以再生出细胞壁并进一步发育成植株。
- 物理特性:原生质体在低渗溶液中会吸水涨破,因此在剥离和培养过程中必须置于等渗或略高渗的溶液环境中。
应试表象与易错陷阱(考点汇总):
- 范围大小判断:对于同一个成熟植物细胞,原生质体 > 原生质层。原生质体比原生质层多出了“细胞核”和“细胞液”这两部分。
- 死细胞的辨别:原生质层表现出选择透过性是以细胞活性为前提的。如果细胞死亡,原生质层会失去选择透过性,变为全透性,此时无法发生质壁分离。
- 质壁分离空间填充物:在质壁分离状态下,细胞壁与原生质层之间的空隙填充的是外界溶液,因为细胞壁是全透性的。
- 实验选材:观察质壁分离通常选用紫色洋葱鳞片叶外表皮,因为它有紫色大液泡,便于观察原生质层(由液泡体积变化推测)的位置变化。
- 原生质滴:这是一个极冷门的干扰项,指精子形成过程中浓缩的细胞质基质,与前两者无直接逻辑关联。
总结来说,原生质体是细胞工程中被剥去外壳的“裸细胞”,承载着遗传和发育的使命;而原生质层是水分代谢中起到屏障作用的“复合膜”,维持着细胞的渗透稳态。
实验的拓展应用:
- 判断细胞死活:只有活细胞的原生质层才具有选择透过性,死细胞不发生质壁分离。
- 测定细胞液浓度范围:通过设置一系列浓度梯度的蔗糖溶液,观察刚发生初始质壁分离时的溶液浓度,即可推测细胞液浓度。
- 特殊溶质的自动复原:若使用一定浓度的 KNO₃、甘油或尿素 溶液,细胞会先发生质壁分离,随后因细胞主动或被动吸收这些溶质,导致细胞液浓度升高,从而发生自动复原。
实验注意事项:
- 浓度控制:蔗糖溶液浓度若过高(如 0.5g/mL),会导致细胞因过度失水而死亡,从而无法观察到复原现象。
- 解离液影响:在观察有丝分裂时使用的解离液(盐酸和酒精)会杀死细胞,因此不能用于质壁分离实验。
这个实验是理解细胞代谢与水分平衡的基础。
人类对通道蛋白的探索历程
人类对通道蛋白的认识经历了从推测到实验证实,再到原子水平结构解析的漫长过程。以下是详细的探索历程:
水通道蛋白的发现:从“间隙扩散”到“专用通道”
早期,科学家普遍认为极小的水分子是穿过磷脂分子的间隙进出细胞的。但后来的一系列观察打破了这一认知:
- 速度矛盾:研究发现,肾脏等部位水分子的跨膜速率远超自由扩散的理论值。
- 同位素证据(1950 年):科学家利用氢同位素标记水分子,发现其通过生物膜的速率明显高于人工膜,暗示了膜上存在特殊“通道”。
- 成功分离(1988 年):美国科学家阿格雷成功分离出了水通道蛋白,确证了其存在。目前已知人类有 13 种水通道蛋白,对肾小球滤过和肾小管重吸收至关重要。

离子通道的探索:从电生理到结构解析
由于离子带有电荷,无法自由穿过脂双层,其通道的发现依赖于电生理技术的突破:
- 早期假说(1960 年代):科学家首先在植物细胞中提出了钾离子通道的概念。
- 工具革命(1976 年):内尔和萨克曼发明了膜片钳技术,使人类能够监测单通道的电流,这一发明让他们获得了 1991 年诺贝尔奖。
- 结构定格(1998 年):麦金农解析了钾离子通道蛋白的立体结构,从分子水平解释了通道如何选择特定离子。
巅峰与未来:诺贝尔奖与医学应用
- 最高荣誉:2003 年,阿格雷(发现水通道)和麦金农(解析离子通道结构)共同获得诺贝尔化学奖。
- 持续研究:目前,科学家已进一步解析了钠、钙等离子的通道结构,并致力于研发调控这些通道的药物,以治疗各种神经和生理疾病。
被动运输(自由扩散和协助扩散)
被动运输是指物质顺浓度梯度(或电化学梯度)进出细胞的过程,此过程不需要消耗细胞代谢产生的能量。它是细胞维持稳态的基本方式之一,主要分为以下两类:
自由扩散 (Simple Diffusion):物质直接通过磷脂双分子层进出细胞。
- 特点:不需要转运蛋白协助,不需要能量。
- 动力:物质两侧的浓度差。浓度差越大,扩散速率越快。
- 实例:气体分子(如 O_2、CO_2、N_2 等)、脂溶性小分子(如甘油、乙醇、苯、性激素等)、极小分子(如水、尿素)。水主要通过协助扩散,但也存在自由扩散。
协助扩散 (Facilitated Diffusion):物质借助细胞膜上的转运蛋白,顺浓度梯度跨膜运输。其特点是需要转运蛋白,不需要能量。运输速率受浓度梯度和转运蛋白数量(饱和性)的限制。
渗透作用 (Osmosis):这是一种特殊的被动运输,专指水分子(或其他溶剂分子)通过半透膜的扩散。
- 方向:从水的相对含量高(溶液浓度低)的一侧向相对含量低(溶液浓度高)的一侧渗透。
- 植物细胞实例:原生质层(细胞膜、液泡膜及两层膜之间的细胞质)相当于半透膜,当外界溶液浓度大于细胞液浓度时,细胞会发生质壁分离。
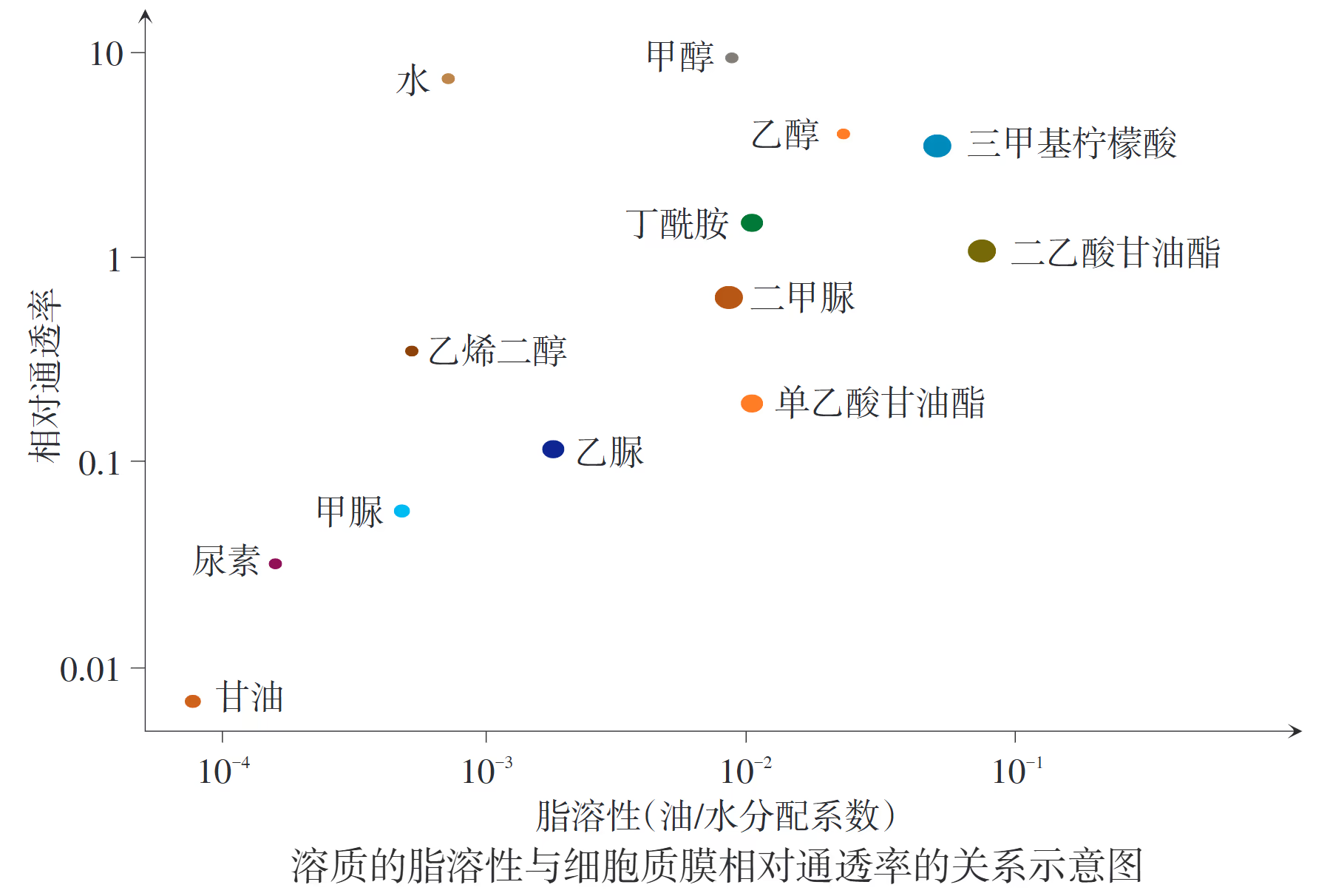
【苏教版】细胞质膜对不同物质有不同的通透性在研究了细胞质膜对许多物质(如水、尿素、甲脲、甘油)的通透性后,科学家发现脂溶性(用油/水分配系数表示,它反映物质在油水两相中的分配情况。系数越大,说明物质越亲油,即脂溶性强;系数越小,越亲水,即水溶性强)越强的物质,越容易通过细胞质膜。
溶质的脂溶性与细胞质膜的相对通透率之间存在显著的正相关关系,即溶质的脂溶性越高,其穿过细胞质膜的相对通透率通常也越高。细胞质膜的基本支架是磷脂双分子层,其内部由磷脂分子的疏水性尾部组成,形成了一个连续的疏水屏障。
- 脂溶性物质(如苯、甘油、性激素、脂溶性维生素 A、D、E、K 等)与磷脂分子具有较强的亲和力,能够直接溶解在脂双层的疏水核心中,通过自由扩散(简单扩散)的方式快速穿过膜。
- 非脂溶性或极性物质(如离子、葡萄糖、氨基酸)则因受疏水尾部的排斥,极难直接通过脂质层,通常需要借助转运蛋白(通道蛋白或载体蛋白)的协助。
1895 年,欧文顿通过上万次实验发现,凡是易溶于脂质的物质,也容易穿过细胞膜;反之,不易溶于脂质的物质,也不容易穿过细胞膜。这一经典发现不仅揭示了脂溶性与通透率的关系,还让科学家推测出细胞膜的主要成分是脂质。
在生物学研究中,溶质的脂溶性常通过油/水分配系数(反映物质在油和水两相中的分配比例)来衡量。
- 系数越大:表示该物质越亲油(脂溶性强),相对通透率越高,如乙醇、三甲基柠檬酸等。
- 系数越小:表示该物质越亲水,通过脂质层的阻力越大,通透率极低。
虽然脂溶性是决定通透率的关键因素,但分子的通透率还受其他物理性质的共同制约:
- 分子大小:在脂溶性相近的情况下,分子越小,穿过磷脂分子间隙的速度越快。
- 电荷与极性:带电荷的离子或强极性分子即使体积很小,其脂溶性也极差,因此对脂质层的通透性极低。
总结:溶质的脂溶性(非极性程度)决定了其在质膜内部疏水环境中的溶解能力。脂溶性越强,物质越容易以简单扩散的方式越过屏障,表现出更高的相对通透率。
转运蛋白(Transport proteins)是镶嵌在细胞膜磷脂双分子层中、负责介导小的有机分子和无机离子进出细胞的膜内在蛋白。根据其转运机制和工作方式的不同,转运蛋白主要分为以下两大类:
载体蛋白 (Carrier Proteins):载体蛋白类似于“旋转式栅门”,通过与特定的溶质分子结合并发生构象改变,将物质从膜的一侧转运到另一侧。
- 工作机制:载体蛋白具有高度的特异性,其结合位点只与特定形状和性质的溶质(如某种离子或葡萄糖)匹配。结合后,蛋白质的构象发生可逆性变化,转运完成后恢复原状。
- 转运速率:相对较慢,每秒通常只能转运 10^3 到 10^6 个分子。
- 功能分类:
- 被动转运(协助扩散):顺浓度梯度转运,不消耗能量,如红细胞上的葡萄糖载体。
- 主动转运(泵):逆浓度梯度转运,需要消耗能量(如 ATP),如钠钾泵 (Na^+-K^+ 泵) 和 Ca^{2+} 泵。
- 协同运输(Cotransport):根据转运方向和数量,可分为单向转运(只运一种溶质)、同向转运(如小肠上皮细胞的葡萄糖-Na^+ 同向转运)和对向转运(如 Na^+-H^+ 交换蛋白)。
通道蛋白 (Channel Proteins):通道蛋白在膜上形成一个贯穿脂双层的亲水性孔道,允许特定大小和电荷的物质顺电化学梯度扩散。
- 工作机制:主要根据分子的大小和电荷进行筛选。物质通过时不需要与蛋白发生紧密结合,且通道蛋白不具有类似载体蛋白的泵功能,因此始终介导被动转运。
- 转运速率:极快,每秒可允许超过 10^6 个离子通过,比载体蛋白快 1000 倍以上。
- 主要类型:
- 水通道蛋白 (Aquaporins):专门转运水分子的通道,广泛存在于肾脏和植物细胞中。
- 离子通道:对特定无机离子(如 Na^+、K^+、Cl^-、Ca^{2+})具有高度选择性。
- 门控机制 (Gating):大多数离子通道是“门控”的,即在受到特定刺激时才瞬间开放:
- 电压门控:响应膜电位的变化(如神经冲动的传导)。
- 配体门控:响应特定化学信号(如神经递质)的结合。
- 压力/应力门控:响应机械力的刺激(如听觉毛细胞受到的震动)。
总结来说,载体蛋白像搬运工,通过变构来回运输;而通道蛋白像隧道,选准对象后让其快速通过。
主动运输(原发继发和光驱动泵)
主动运输是细胞为了维持生命活动,通过载体蛋白的协助,消耗代谢能量(通常是 ATP),将物质逆浓度梯度(或逆电化学梯度)跨膜转运的过程。
以下是主动运输的详细分类与作用机制:
原发性主动运输(直接消耗 ATP):这类运输直接利用 ATP 水解释放的能量来驱动物质“上坡”运输,相关的载体蛋白通常被称为“泵”。
- 钠钾泵 (Na^+-K^+ 泵):这是动物细胞中最典型的例子。它每消耗 1 分子 ATP,就会向细胞外泵出 3 个 Na^+,向细胞内泵入 2 个 K^+。这维持了细胞内高钾、细胞外高钠的状态,对产生神经冲动和维持渗透压至关重要。
- 钙泵 (Ca^{2+} 泵):负责将细胞质基质中的 Ca^{2+} 泵出细胞或泵入内质网,维持胞内极低的钙离子浓度。
- 质子泵 (H^+ 泵):普遍存在于植物、真菌和细菌的质膜上,通过泵出 H^+ 创造跨膜的质子浓度梯度。
继发性主动运输(间接消耗能量/协同运输):这类运输不直接消耗 ATP,而是利用原发性主动运输建立的离子电化学梯度(通常是 Na^+ 或 H^+ 梯度)作为动力,带动另一种物质逆浓度梯度运输。
- 同向转运 (Symport):被转运物质与动力离子向同一方向运动。例如,小肠上皮细胞利用 Na^+ 的向内流动的动力,将葡萄糖逆浓度梯度吸入体内。
- 对向转运 (Antiport):被转运物质与动力离子向相反方向运动。例如,Na^+-H^+ 交换蛋白利用 Na^+ 进入细胞的动力将 H^+ 排出,以此调节细胞内的 pH 值。
光驱动泵:主要存在于某些细菌(如盐杆菌)中,利用光能直接驱动物质的主动运输,例如细菌紫膜质利用阳光将质子泵出细胞。
主动运输的生物学意义:
- 维持稳态:确保细胞内部某些离子或分子的浓度显著高于或低于外界环境。
- 高效吸收:允许细胞从环境浓度极低的情况下,依然能主动摄取必需的营养物质(如氨基酸、葡萄糖、无机盐)。
- 排除废物:将细胞内代谢产生的有害物质或废物逆浓度排出。
主动运输的速率通常受到载体蛋白数量和能量供应(如氧气浓度、温度影响呼吸作用)的限制。
区分:继发性主动运输和协助扩散,有的时候看起来很类似(载体本身都不直接消耗 ATP 或光能)。但实际上,判断的唯一标准是看该载体蛋白运输的所有物质的顺逆浓度梯度情况。只有当一个载体蛋白运输的所有物质都是顺浓度梯度,且没有额外能量(ATP/光能)供应时,这个整体过程才是协助扩散;只要运输过程中,包含有逆浓度梯度的物质,这整个过程就一定是主动运输。哪怕载体蛋白没有直接消耗 ATP,它也会利用“另一种物质顺浓度梯度释放的势能”(这就是继发性主动运输)。归根结底,用来提供势能的浓度梯度是由其他耗能泵维护的。只要系统最终消耗了能量来实现逆梯度转运,本质就是主动运输。
胞吞与胞吐
胞吞(Endocytosis)与胞吐(Exocytosis)是细胞处理蛋白质、多糖以及大型颗粒等大分子物质的特殊方式。由于这些物质体积巨大,无法直接通过跨膜运输(如自由扩散或主动运输)进入细胞,必须依赖膜的变形来实现转运。
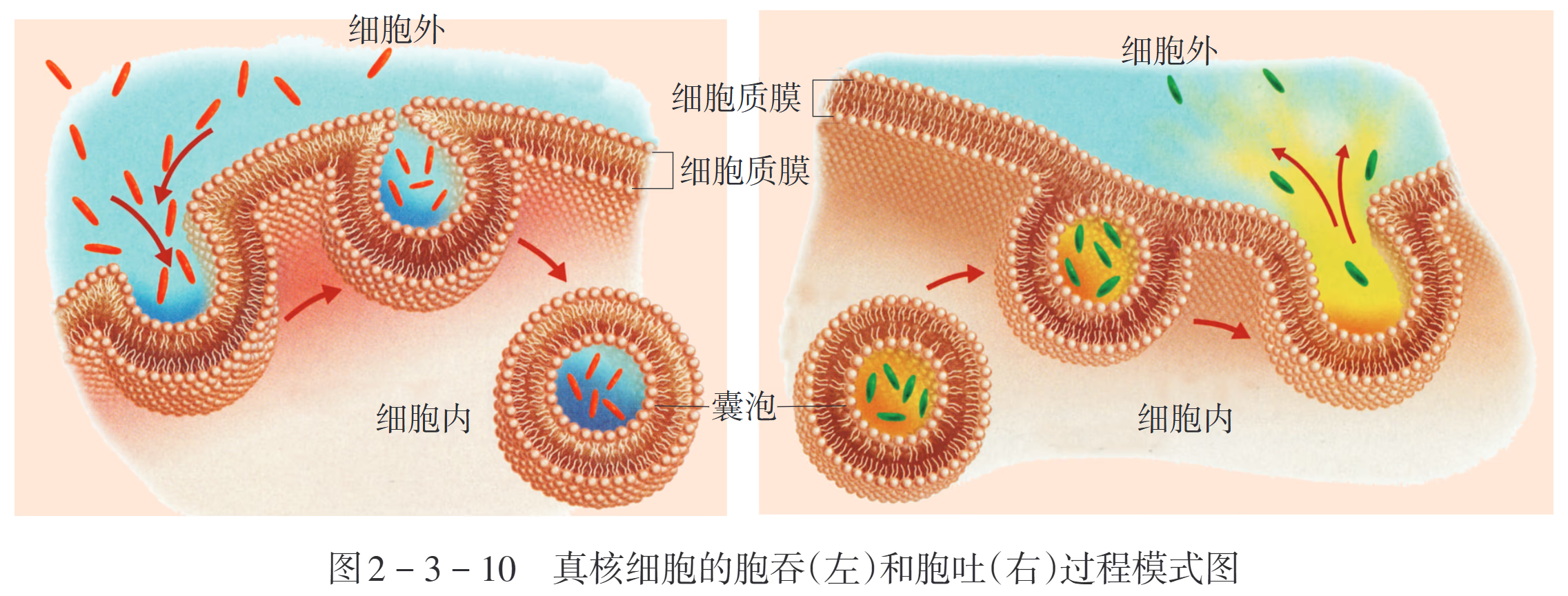
胞吞作用是细胞通过质膜内陷包裹外界物质形成囊泡,并将其带入细胞的过程。根据摄取物质的物理状态和机制,可分为以下三类:
- 吞噬作用(Phagocytosis,即“细胞吃东西”):细胞伸出伪足包围大型固体颗粒(如细菌、细胞碎片),形成吞噬体(Phagosome)。例如,人体内的中性粒细胞和巨噬细胞能通过这种方式消灭侵入的病原体。
- 胞饮作用(Pinocytosis,即“细胞喝东西”):细胞非特异性地摄入细胞外液及其中溶解的小分子溶质,形成微小的胞饮泡。这是一个连续不断的过程,有助于细胞调节质膜面积和容积。
- 受体介导的胞吞作用(Receptor-mediated Endocytosis):这是一种极具专一性的高效摄取方式。目标分子(配体)与质膜上的特异性受体结合,诱导网格蛋白(Clathrin)在膜内侧聚集并形成包被小窝,最终脱离形成包被囊泡。人体吸收胆固醇(通过低密度脂蛋白 LDL 颗粒)就是典型的例子。
胞吐作用是细胞将胞内的大分子物质(如分泌蛋白、神经递质、废物)通过囊泡运送到质膜,并与之融合释放到细胞外的过程:
- 组成型胞吐:所有真核细胞中持续进行的“默认途径”,用于更新质膜蛋白和脂质,或分泌胞外基质组分。
- 调节型胞吐:仅存在于特化的分泌细胞中。物质(如胰岛素、消化酶)储存在分泌囊泡内,直到接收到特定的胞外信号(如血糖升高)后,才触发囊泡与质膜融合释放。
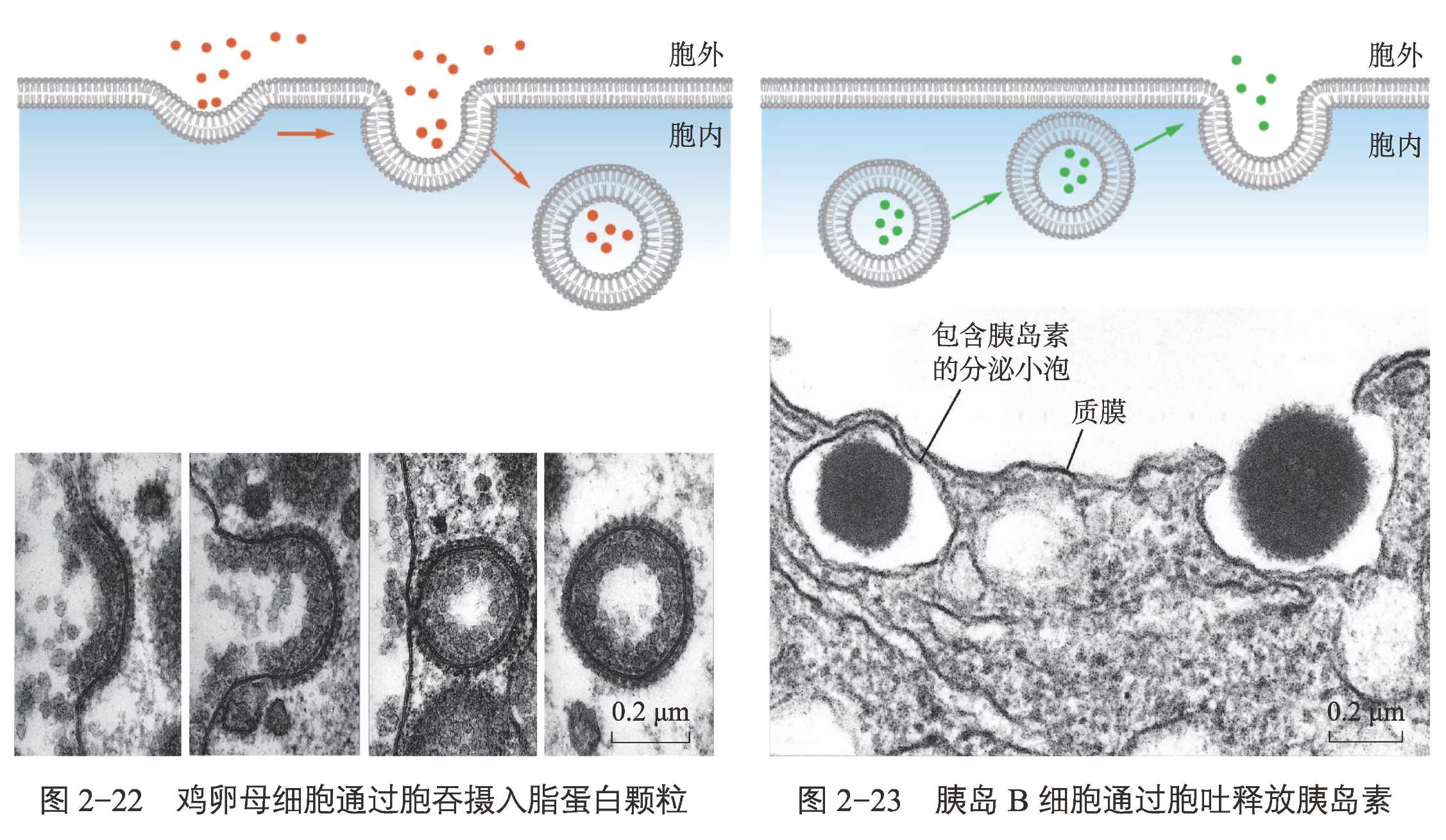
结构基础与能量需求:
- 膜的流动性(Fluidity):胞吞和胞吐的实现核心在于细胞膜的磷脂双分子层和蛋白质不是静止的,而是具有一定流动性的。膜的断裂与融合正是基于这一物理特性。
- 能量消耗:这两种方式均属于耗能过程,需要细胞呼吸产生的 ATP 提供动力。
为什么分泌蛋白必须由附着在内质网上的核糖体合成?这是由蛋白质的分选信号和转运机制决定的:
- 核糖体分工:游离核糖体合成的蛋白质通常留在细胞溶胶中发挥作用(如糖酵解酶)。而分泌蛋白、膜蛋白及溶酶体酶则由附着在内质网上的核糖体合成。
- 信号肽(Signal Peptide)的作用:合成开始时,肽链 N 端的信号肽会被信号识别颗粒(SRP)识别,引导整个核糖体附着到内质网上。随后,肽链直接穿入内质网腔内进行初步加工。
- 囊泡运输路径:进入内质网的蛋白质被包装在囊泡中,依次经过高尔基体进行修饰、分拣和包装,最终通过胞吐排出细胞。由于蛋白质分子巨大且具亲水性,它们无法通过自由扩散离开细胞溶胶,必须在膜性区室(内质网 - 高尔基体)中“接力”转运,以保证其不与细胞溶胶成分混合。
典型案例:痢疾内变形虫的攻击手段。痢疾内变形虫充分利用了这两种机制。它通过胞吐分泌蛋白分解酶来溶解人体的肠壁组织,造成溃疡;随后通过胞吞作用(吞噬)吃掉受损的肠壁细胞和红细胞,导致严重的阿米巴痢疾。
医疗:人工膜
人工膜(Artificial membrane)是指利用混合脂质或一种(或少数几种)纯磷脂,在实验室条件下人工制备的各种模拟生物膜结构。它是体外研究生物膜结构与功能、膜蛋白与膜脂相互作用的重要手段。
人工膜的构建基于脂类分子的两亲性(亲水头部和疏水尾部),在水环境中会自发聚集以减少能量不稳定性。
- 脂质体(Liposome):这是最常见的人工膜形式。当磷脂分子在水中受到超超声波处理或搅拌时,会自发形成充满液体的球状小泡。
- 结构:脂质体由磷脂双分子层组成,疏水尾部相对位于内侧,亲水头部朝向内外的水溶液环境。这使得脂质体内部有一个水溶液环境,而两层磷脂分子之间则是一个疏水区。
- 单分子层膜与累积膜:脂质在空气 - 水界面上可以铺展成单分子层(Monolayer)。若将玻片垂直于该界面上下移动,可形成达数百层厚的人工膜,称为累积膜。
- 黑膜(Black Lipid Membrane):在两侧都是水溶液的隔板小孔上形成的平坦磷脂双分子层。因其在光照下因干涉消失而显黑色,故名黑膜,常用于电生理研究。
- LB 膜(Langmuir-Blodgett membrane):由一个或多个单层有机材料组成的精细人工膜,常用于体外膜脂和膜蛋白的相关研究。
由于天然生物膜成分极其复杂(含有多种脂质、蛋白质和糖类),研究单一组分的功能非常困难。
- 简化模型研究:科学家将纯化的膜蛋白(如离子通道或载体蛋白)重新嵌入人工脂质体中(这一过程称为生物膜的重建),可以在比天然膜更简单、受控的环境中研究特定蛋白质的转运功能或酶活性。
- 验证膜特性:通过对比“人工脂质双分子层”和“天然生物膜”对物质的通透性,科学家证明了水分子、气体分子等可以直接穿过脂双层,而离子和极性大分子(如葡萄糖)则必须依赖膜上的特定转运蛋白。
- 合成生物学应用:人工膜可用于构建“人造细胞”。例如,将细菌视紫红质和 ATP 合酶合并到巨大的单膜小泡中,构建出能利用光能驱动内部 DNA 转录翻译的人造细胞。
人工膜技术不仅停留在理论研究,已广泛应用于临床和工业领域:
- 药物运载(“生物导弹”):脂质体可作为药物或 DNA 的载体。
- 载药机制:水溶性药物包裹在脂质体内部的水溶液环境中,脂溶性药物则包在两层磷脂分子之间的疏水层中。
- 靶向性:在脂质体表面镶嵌特异性抗体或信号分子,可以使其特异性地结合到靶细胞(如癌细胞)表面,通过膜融合或胞吞作用将药物送入细胞,减少对健康组织的损伤。
- 人工器官(透析型人工肾):当肾功能发生障碍导致代谢废物(如尿素)无法排出时,临床常用透析型人工肾。
- 原理:其核心部件是人工合成的血液透析膜(一种人工膜材料),利用了生物膜的选择透过性原理。血液流经人工肾时,透析膜能将血液中的代谢废物和多余水分透析掉,而保留血细胞和大分子蛋白质。
- 工业分离与环境治理:
- 海水淡化:模拟细胞膜的选择透过性,利用人工膜滤去海水中的盐分。
- 污水处理:有选择地阻挡有毒重金属离子,降低水污染。
与天然生物膜的区别(易混易错点):
生物活性:天然生物膜具有生物活性,其选择透过性依赖于活细胞的代谢和转运蛋白。
选择性差异:仅含脂质成分的人工膜属于半透膜,其通透性主要取决于溶质分子的大小和极性(脂溶性);而含有特定蛋白质的天然生物膜或重建人工膜才具备高度的选择透过性。