酶、代谢与呼吸作用
能量概述
酶的探究历程
这段关于“酶”的探究历程,生动地展示了科学认知是如何通过观察、实验、争论和技术突破而不断演进的。以下是根据你提供的纲要进行的详细展开:
早期探索,从物理消化到化学消化的转折:
- 斯帕兰札尼的实验(1773 年):在 18 世纪之前,人们普遍认为胃只能通过物理摩擦来磨碎食物。斯帕兰札尼将肉块放入带孔的小金属笼内让鹰吞下,这一设计的精妙之处在于笼子能排除胃物理摩擦的影响,但允许胃液进入。肉块的消失证明了胃液中存在某种能分解有机物的化学物质。
- 施旺发现胃蛋白酶(1835 年):德国科学家施旺在研究胃腺分泌物时,发现将其与盐酸混合后,对肉类的分解能力远超盐酸单干。他成功提取出了这种有效成分,并命名为胃蛋白酶,这是人类历史上第一次从动物体内分离出酶。
核心概念:细胞代谢离不开酶:
- 细胞代谢定义:细胞内部每时每刻都在进行的成千上万种化学反应统称为细胞代谢,它是生命活动的基础。
- 酶的必要性:许多化学反应在体外需要高温、高压、强酸或强碱等剧烈条件,但细胞内环境温和且稳定(常温、常压、pH 接近中性)。在这种环境下,只有靠酶的催化作用,代谢才能高效、有序地进行。
- 案例:过氧化氢(\ce{H2O2})的分解:\ce{H2O2} 是代谢产生的有害废物。细胞内的过氧化氢酶能极快地将其分解为水和氧气,单分子催化速率可达每秒 4000 万个分子。相比无机催化剂(如 \ce{Fe^3+}),酶的催化效率高出 10^7 \sim 10^{13} 倍,本质在于酶能显著降低反应的活化能。
酶本质的深度探索历程:
酿酒中的争论(19 世纪):
巴斯德(1857 年):认为发酵是活酵母细胞代谢的结果,没有活细胞就不能发酵。
李比希:认为发酵只是纯化学反应,只需要酵母细胞死亡裂解后释放的某些物质参与即可。
毕希纳与“酿酶”(1897 年):毕希纳用石英砂研磨酵母细胞并加压过滤,发现无细胞提取液也能使葡萄糖产生酒精。这证明了引起发酵的是细胞内的物质(他称为酿酶),而非细胞生命活动本身,从而终结了上述争论。
萨姆纳证明酶是蛋白质(1926 年):在很长一段时间内,酶被认为是不解之谜。萨姆纳坚信酶是蛋白质,历时 9 年从刀豆中提取出脲酶结晶,并证明其本质。随后,胃蛋白酶、胰蛋白酶等结晶的获得确认了“绝大多数酶是蛋白质”这一共识。
核酶的发现(20 世纪 80 年代):切赫和奥尔特曼发现少数 RNA 也具有催化功能(核酶),这一发现突破了酶只能是蛋白质的传统观念,开辟了生化研究的新领域。
讨论与总结:科学方法论。
- 观点评述:巴斯德强调了生物体与代谢的联系,李比希关注化学本质;虽然各具局限性,但他们的争论聚焦了研究目标。
- 争论的意义:科学争论能促使研究者不断设计实验去证伪或证实,推动认知走向本质。
- 毕希纳的贡献:证明了酶可以脱离细胞在体外发挥催化作用。
- 萨姆纳的精神:体现了不畏艰辛、持之以恒、实事求是的科学态度。
- 酶的完整定义:酶是活细胞产生的具有催化作用的有机物,其中绝大多数是蛋白质,少数是 RNA。
酶的探究实验
这三个实验是高中生物学中关于酶的特性及其作用原理的核心探究活动。以下是根据提供的资料进行的详细展开讲解:
比较过氧化氢在不同条件下的分解:该实验旨在通过观察过氧化氢(\ce{H2O2})在不同条件下的分解速率,了解酶的作用原理及高效性。
- 实验原理:\ce{H2O2} 在常温下分解极慢,但在加热、无机催化剂(如 \ce{Fe^3+})或新鲜肝脏研磨液(含过氧化氢酶)的作用下会加速分解产生 \ce{O2}。可以通过气泡产生的多少和带火星的卫生香复燃的剧烈程度来判断反应速率。
- 实验步骤与预期现象:
- 1 号试管(对照组):常温处理,几乎无气泡产生,卫生香不复燃。
- 2 号试管(受热):\pu{90^oC} 水浴加热,产生少量气泡,卫生香不复燃。这说明加热能为反应提供能量,提高反应速率。
- 3 号试管(无机催化剂):滴加 2 滴 3.5\% \ce{FeCl3} 溶液,产生较多气泡,卫生香复燃但不明显。
- 4 号试管(过氧化氢酶):滴加 2 滴 20\% 新鲜肝脏研磨液,产生大量气泡,卫生香迅速复燃。
- 实验结论:
- 酶具有催化作用。
- 高效性:酶的催化效率远高于无机催化剂(约 10^7 \sim 10^{13} 倍),其本质是酶降低化学反应活化能的作用更显著。
- 注意事项:必须使用新鲜的肝脏研磨液,否则过氧化氢酶会被微生物分解或因放置时间过长而失活。
淀粉酶对淀粉和蔗糖的水解作用:该实验用于证明酶具有专一性,即一种酶只能催化一种或一类化学反应。
- 实验原理:淀粉和蔗糖是非还原糖,但它们在相应酶的作用下水解产生的麦芽糖、葡萄糖、果糖等属于还原糖。还原糖与斐林试剂在水浴加热条件下会产生砖红色沉淀。
- 实验设计与步骤:
- 1 号试管:注入 \pu{2 mL} 淀粉溶液 + \pu{2 mL} 淀粉酶溶液。
- 2 号试管:注入 \pu{2 mL} 蔗糖溶液 + \pu{2 mL} 淀粉酶溶液。
- 将两管放入 \pu{60 ^oC}(或 \pu{37 ^oC})温水中保温 5 分钟,然后加入斐林试剂并水浴加热。
- 实验现象:1 号试管出现砖红色沉淀,2 号试管保持蓝色(斐林试剂本色)。
- 结论:淀粉酶只能催化淀粉水解,不能催化蔗糖水解,证明酶具有专一性。
- 关键提示:不宜选用碘液作为检测试剂,因为碘液只能检测淀粉是否被分解,无法检测蔗糖是否被水解。
影响酶活性的条件:该实验探究温度和 pH 对酶活性的影响,体现了酶作用条件较温和的特性。
- 探究温度对酶活性的影响:
- 材料选择:通常用淀粉酶。不能使用过氧化氢酶,因为 \ce{H2O2} 本身受热会加速分解,干扰实验结果。
- 方法:设置 \pu{0 ^oC}、约 \pu{60 ^oC} 和 \pu{100 ^oC}(沸水)三组温度。先将底物和酶分别在相应温度下保温后再混合。
- 检测:使用碘液。在最适温度下淀粉被完全水解,滴加碘液不变蓝;在高温或低温下淀粉未完全水解,滴加碘液变蓝。
- 探究 pH 对酶活性的影响:
- 材料选择:常用过氧化氢酶。
- 方法:设置酸性(\ce{HCl})、中性(蒸馏水)和碱性(\ce{NaOH})三个梯度的溶液环境。
- 现象:在中性(或最适 pH)环境下,产生气泡最多;在强酸或强碱环境下,基本无气泡产生。
- 核心结论:
- 酶在最适温度和最适 pH 下活性最高。
- 失活与抑制:高温、过酸、过碱会破坏酶的空间结构,导致酶永久失活;而低温会抑制酶的活性,但空间结构稳定,在适宜温度下活性可恢复。
这些实验加深了我们对酶作为生物催化剂本质的理解。
课本:酶的基本概念
能量的释放、储存和利用,都必须通过化学反应来实现。细胞中每时每刻都进行着许多化学反应,统称为细胞代谢(cellular metabolism)。细胞代谢离不开酶(enzyme)。
加热能促进过氧化氢分解,是因为加热使过氧化氢分子得到了能量,从常态转变为容易分解的活跃状态。分子从常态转变为容易发生化学反应的活跃状态所需要的能量称为活化能(activation energy)。
\ce{Fe^3+} 和过氧化氢酶能促进过氧化氢分解,它们没有给过氧化氢分子提供能量,而是降低了过氧化氢分解反应的活化能。如果把化学反应比作驾车翻越一座高山,“加热”相当于给汽车加大油门,用催化剂则相当于帮司机找到穿山的隧道。与无机催化剂相比,酶降低活化能的作用更显著,催化效率更高。正是由于酶的催化作用,细胞代谢才能在温和条件下快速有序地进行。
酶的化学本质不同于无机催化剂。一般来说,酶是活细胞产生的具有催化作用的有机物,其中绝大多数酶是蛋白质。酶的催化作用与无机催化剂有什么不同呢?
酶具有高效性。大量的实验数据表明,酶的催化效率是无机催化剂的 107 ~ 1013 倍。无机催化剂催化的化学反应范围比较广。例如,酸既能催化蛋白质水解,也能催化脂肪水解,还能催化淀粉水解。酶能像无机催化剂一样,催化多种化学反应吗?目前已发现的酶有 8 000 多种,它们分别催化不同的化学反应。
细胞中几乎所有的化学反应都是由酶催化的。酶催化特定化学反应的能力称为酶活性(enzyme activity)。酶活性可用在一定条件下酶所催化某一化学反应的速率表示。
实验过程中的变化因素称为变量。其中人为控制的对实验对象进行处理的因素叫作自变量,上述实验中加热、加 \ce{FeCl3} 溶液、加肝脏研磨液,是对过氧化氢溶液的不同处理,温度和催化剂都属于自变量;因自变量改变而变化的变量叫作因变量,上述实验中过氧化氢分解速率就是因变量。除自变量外,实验过程中还存在一些对实验结果造成影响的可变因素,叫作无关变量。如上述实验中反应物的浓度和反应时间等。除作为自变量的因素外,其余因素(无关变量)都保持一致,并将结果进行比较的实验叫作对照实验。对照实验一般要设置对照组和实验组,上述实验中的 1 号试管就是对照组,2 号、3 号和 4 号试管是实验组。本实验的对照组未作任何处理,这样的对照组叫作空白对照。想一想,设置这样的对照组有什么意义?
过氧化氢酶只能催化过氧化氢分解,不能催化其他化学反应。脲酶除了催化尿素分解,对其他化学反应也不起作用。每一种酶只能催化一种或一类化学反应。细胞代谢能够有条不紊地进行,与酶的专一性是分不开的。许多无机催化剂能在高温、高压、强酸或强碱条件下催化化学反应。
溶液的温度和 pH 都对酶活性有影响。与无机催化剂相比,酶所催化的化学反应一般是在比较温和的条件下进行的。科学家采用定量分析的方法,分别在不同的温度和 pH 条件下测定同一种酶的活性,根据所得到的数据绘制成曲线图。分析这两个曲线图可以看出,在最适宜的温度和 pH 条件下,酶的活性最高。温度和 pH 偏高或偏低,酶活性都会明显降低。
一般来说,动物体内的酶最适温度在 \pu{35 \sim 40 ^oC} ;植物体内的酶最适温度在 \pu{40 \sim 50 ^oC} ;细菌和真菌体内的酶最适温度差别较大,有的酶最适温度可高达 \pu{70 ^oC}。动物体内的酶最适 pH 大多在 6.5 \sim 8.0,但也有例外,如胃蛋白酶的最适 pH 为 1.5;植物体内的酶最适 pH 大多为 4.5 \sim 6.5。
过酸、过碱或温度过高,会使酶的空间结构遭到破坏,使酶永久失活。在 \pu{0 ^oC} 左右时,酶的活性很低,但酶的空间结构稳定,在适宜的温度下酶的活性会升高。因此,酶制剂适宜在低温下保存。
20 世纪 60 年代以前,医院里用的葡萄糖是用盐酸催化淀粉水解的方法来生产的,生产过程需要在 \pu{245 kPa} 的高压和 \pu{140 \sim 150 ^oC} 的高温下进行,并且需要耐酸的设备。60 年代以后改用酶法生产。
细胞中的各类化学反应之所以能有序进行,还与酶在细胞中的分布有关。拿植物叶肉细胞来说,与光合作用有关的酶分布在叶绿体内,与呼吸作用有关的酶分布在细胞质基质和线粒体内,这样,光合作用与呼吸作用在细胞内不同的区室同时进行,就可以互不干扰了。
酶的应用
这些课本中提到的酶的应用,展示了酶作为生物催化剂如何通过其高效性和专一性,在医疗、工业和环保等领域发挥巨大作用。以下是详细的展开讲解:
医疗卫生,抗菌、助消化与溶栓:
- 溶菌酶(Lysozyme):这种酶能特异性地水解细菌细胞壁中肽聚糖的化学键(NAG 与 NAM 之间的糖苷键),使细菌吸水破裂死亡。在临床上,它常与抗生素联用产生协同效应,增强杀菌力。
- 助消化药物:多酶片包含胃蛋白酶、胰淀粉酶、胰脂肪酶和胰蛋白酶等。它们能帮助将食物中的蛋白质、淀粉和脂肪分解为可吸收的小分子,用于治疗消化不良。
- 创伤处理与溶栓:胰蛋白酶能去除伤口坏死组织,促进愈合。针对心肌梗死等血栓疾病,医生会使用组织型纤溶酶原激活剂(t-PA)或尿激酶,它们能激活纤溶酶原转化为纤溶酶,从而溶解血块。
工业生产,提升效率与品质:
- 食品加工:
- 果胶酶:能分解果肉细胞壁中的果胶,显著提高果汁的出汁率并使果汁更清亮。
- 凝乳酶(Chymosin):在奶酪生产中,它能使牛奶中的酪蛋白凝集,最初从牛胃提取,现在已能通过转基因微生物大规模生产。
- 日化用品:加酶洗衣粉中添加了蛋白酶、脂肪酶和淀粉酶。为了使其在漂白剂存在下仍能保持活性,科学家利用蛋白质工程将枯草杆菌蛋白酶(Subtilisin)中易被氧化的氨基酸(如 222 位甲硫氨酸)替换,从而显著提升了酶的稳定性。
环境保护与绿色能源:
- 废物处理:脂肪酶能催化废弃油脂转化成生物柴油,实现资源合理利用并保护环境。
- 造纸与降解:在造纸工业中,利用酶替代化学法进行木质素降解和纸张漂白,不仅降低成本,还减少了环境污染。
酶工程对自然的改造。自然界中存在的酶有时在高温、强酸碱或有机溶剂下容易失活:
- 稳定性改造:例如利用定点突变技术在 T4 溶菌酶中引入二硫键,显著提高了其耐热性。
- 抗药性改造:青霉素酰化酶可以将天然青霉素改造成氨苄青霉素等杀菌力更强、广谱性更好的半合成抗生素。
酶的应用是否让你感受到了生物技术的奇妙?
酶抑制剂概述
酶抑制剂是指能够与酶分子结合并降低其催化活性的化学物质,它们在调节细胞代谢、药物开发及农业生产中具有核心地位。根据抑制剂与酶结合的性质及动力学特征,通常将其分为以下类别:
可逆抑制剂(Reversible Inhibitors):此类抑制剂通过非共价键与酶结合,抑制作用是暂时的,可以通过物理方法(如透析)移除。
竞争性抑制剂(Competitive Inhibitor):
- 机制:其分子结构通常与底物非常相似,因此能与底物竞争酶的活性位点。
- 特征:这种抑制是可逆的,增加底物浓度可以减弱甚至消除抑制作用。
- 动力学参数:最大反应速率(V_{max})保持不变,但米氏常数(K_m)增大(意味着酶与底物的表观亲和力下降)。
- 典型案例:磺胺类药物竞争性抑制细菌叶酸合成酶;丙二酸抑制琥珀酸脱氢酶。
非竞争性抑制剂(Non-competitive Inhibitor):
- 机制:抑制剂结合在酶活性位点之外的别构位点(Allosteric site),引起酶的空间结构改变。
- 特征:这种构象变化使得底物无法再与活性中心契合,或使已结合的 ES 复合物无法生成产物,因此无法通过增加底物浓度来解除。
- 动力学参数:V_{max} 减小,K_m 不变。
- 典型案例:重金属离子(如汞、铅)通过结合酶的侧链基团使酶失活。
反竞争性抑制剂(Uncompetitive Inhibitor):
- 机制:抑制剂仅与“酶 - 底物复合物”(ES)结合,而不结合游离酶。
- 动力学参数:V_{max} 和 K_m 同时减小。
不可逆抑制剂(Irreversible Inhibitors):这类抑制剂通常与酶活性中心的必需基团形成牢固的共价键,导致酶永久失活。
- 特征:抑制程度随时间增加而加强,无法通过物理方法除去,效果相当于降低了系统中有效酶的浓度。
- 典型案例:
- 有机磷农药/神经毒气:特异性结合乙酰胆碱酯酶的丝氨酸残基,导致神经系统信号传导中断。
- 青霉素:不可逆地抑制细菌合成细胞壁的关键酶,使细菌吸水破裂死亡。
在细胞代谢中,酶抑制剂不仅是外源药物,也是天然的调节工具。
- 机制:代谢途径的终产物(End product)往往作为抑制剂(通常是别构抑制),作用于该途径起始阶段的一个或多个酶。
- 意义:这是一种精密的负反馈调节(反馈抑制),能防止产物过量积累,确保细胞代谢有序进行。
许多现代药物本质上就是特定酶的抑制剂:
- 他汀类药物(Statins):通过抑制 HMG-CoA 还原酶来降低胆固醇水平。
- 格列卫(Gleevec):特异性结合超活化激酶的 ATP 结合区,阻断导致白血病的癌症信号。
这些抑制机制展示了生物分子之间精妙的契合与调节。
呼吸作用
新陈代谢
在深入探讨新陈代谢(Metabolism)这一生命最基本的特征时,我们需要从分子逻辑、细胞机制以及人体系统协调这三个层面进行重构。新陈代谢不仅是生物体内所有化学反应的总称,更是生命系统维持高度有序状态、抵御熵增(无序化)的核心手段。
在细胞水平上,新陈代谢由两套方向相反、但又精密耦合的化学反应组成:合成代谢(同化作用)与分解代谢(异化作用)。
- 分解代谢(Catabolism):这是一个“降解”的过程。细胞将复杂的有机大分子(如糖类、脂肪、蛋白质)分解为更简单的分子(如二氧化碳、水、尿素等)。
- 核心功能:释放储存在化学键中的能量,并将其捕获到活化载体分子(主要是 \ce{ATP})中;同时产生合成反应所需的“结构单元”(小分子)。
- 合成代谢(Anabolism):这是一个“构建”的过程。细胞利用分解代谢提供的能量和原料,合成自身所需的蛋白质、核酸、多糖等复杂大分子。
- 热力学本质:生命是一个开放的“耗散结构”。由于合成反应是熵减的过程(有序性增加),它必须与分解反应释放的能量偶联,并向环境排出热量(熵增),以符合热力学第二定律。
细胞内几乎所有的需能过程都不直接使用葡萄糖,而是通过 \ce{ATP}(腺苷三磷酸) 这一能量货币。
- 结构特性:\ce{ATP} 含有两个不稳定的高能磷酸键(磷酐键),末端磷酸基团具有较高的转移势能。
- 能量偶联:细胞通过 \ce{ATP} 的水解(释放能量)来驱动吸能反应(如主动运输、肌肉收缩、物质合成);同时通过细胞呼吸(放能反应)将 \ce{ADP} 和磷酸重新合成 \ce{ATP}。
细胞代谢之所以能在常温、常压的温和环境下高效、有序进行,完全依赖于酶(Enzymes)。
- 作用机理:酶通过显著降低化学反应所需的活化能,使原本极其缓慢的反应在瞬间完成。
- 特性:具有极高的催化效率、专一性(一酶催一反应)以及对环境(pH、温度)的高度敏感性。
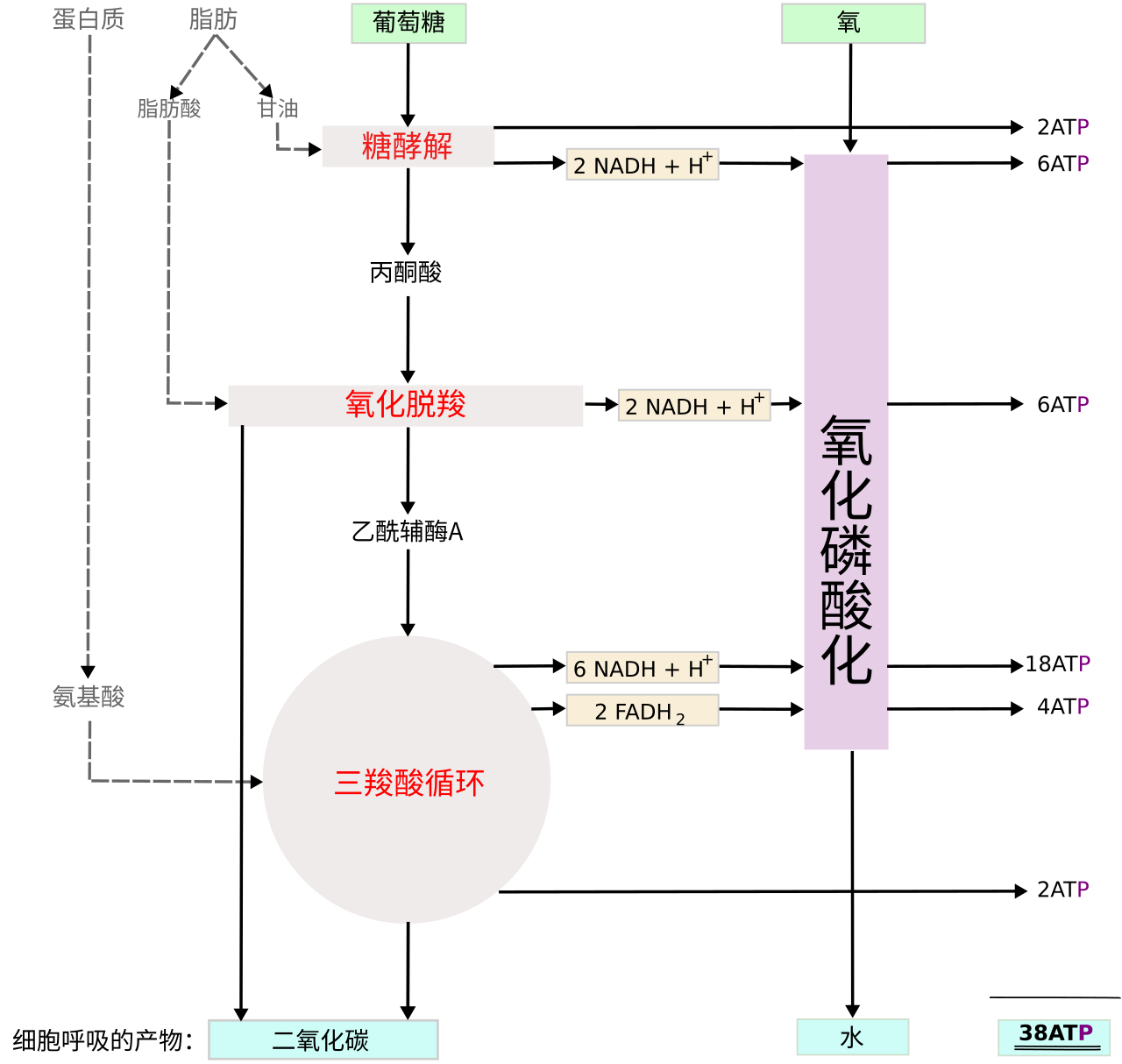
细胞呼吸是细胞获取能量的主要途径,主要分为有氧呼吸和无氧呼吸。
- 有氧呼吸(主要途径):在氧气参与下,葡萄糖被彻底氧化分解为 \ce{CO2} 和 \ce{H2O}。
- 场所:始于细胞质基质(糖酵解),完成于线粒体(柠檬酸循环与电子传递链)。
- 产能:1 分子葡萄糖彻底氧化可产生约 32 个 \ce{ATP}。
- 无氧呼吸:在缺氧条件下,葡萄糖不完全分解产生乳酸或酒精,释放少量能量。人体在剧烈运动时,肌肉细胞会进行乳酸发酵以提供紧急能量。
人体的新陈代谢不是细胞活动的简单加和,而是由多个器官系统协同完成的物质与能量流转过程。物质摄取与废物排出的链条:
- 消化系统:将食物大分子分解为单糖、氨基酸、脂肪酸,这是代谢的第一阶段(消化阶段)。
- 呼吸系统:为细胞提供电子最终受体(\ce{O2}),并排出代谢废气(\ce{CO2})。
- 循环系统:充当“物流网络”,将营养物质和氧气运送至全身组织细胞,同时将代谢废物运往排泄器官。
- 泌尿系统与皮肤:负责排出大部分含氮废物(如尿素)、多余的水和无机盐,维持内环境的水盐平衡。
代谢调节与内环境稳态(Homeostasis):人体内环境(血浆、组织液等)的理化性质(如血糖浓度、温度、pH)必须保持相对稳定,新陈代谢才能正常进行。
- 器官分工:
- 肝脏:代谢的中心实验室,调节血糖浓度,将有毒的氨转化为尿素。
- 肌肉:产热的主要来源(战栗产热与运动产热),高强度代谢消耗大量 \ce{ATP}。
- 神经网络调节:
- 体液调节:二氧化碳浓度升高会刺激化学感受器,调节呼吸运动的深度和频率。
- 激素调节:胰岛素和胰高血糖素通过拮抗作用维持血糖稳态。甲状腺激素则能提高细胞代谢的速率,促进生长发育。
如果新陈代谢或其调节机制出现障碍,会导致一系列疾病。例如,肾功能衰竭(尿毒症)会导致代谢废物在内环境积聚,引起全身性中毒;糖尿病则是由于糖代谢调节受损导致的稳态失调。
新陈代谢是生命在微观分子逻辑上的高度精准耦合(由酶和 \ce{ATP} 驱动)与在宏观系统层面的高度协调一致(由神经-体液-免疫调节维持)的统一体。它不仅是生物体生长、增殖的基础,更是生命之所以能成为“活”的系统的根本原因。
能量货币 ATP
三磷酸腺苷(\ce{ATP})被形象地称为细胞的“能量货币”,是生物体内最直接、最重要的能量来源。\ce{ATP} 的化学本质是一种核苷酸,由三个部分组成:
- 腺嘌呤(Adenine):一个含氮碱基。
- 核糖(Ribose):一个五碳糖,与腺嘌呤结合形成腺苷(Adenosine)。
- 磷酸基团(Phosphate groups):三个磷酸基团依次连接在核糖上,分别称为 \alpha、\beta 和 \gamma 磷酸。
“高能”的秘密,磷酸酐键。\ce{ATP} 分子中连接磷酸基团的化学键被称为磷酸酐键(常俗称为“高能磷酸键”,用“~”表示):
- 不稳定性:由于三个磷酸基团在生理 pH 下都带有负电荷,它们之间存在强烈的静电排斥力,这使得 \ce{ATP} 分子极不稳定。
- 能量释放:当末端的磷酸键断裂(水解)时,会释放出大量的自由能。在活细胞内,1 mol \ce{ATP} 水解为 \ce{ADP} 和 \ce{Pi} 释放的能量约为 \pu{57 kJ/mol},几乎是标准条件下(\pu{30.5 kJ/mol})的两倍。
\ce{ATP}-\ce{ADP} 循环,细胞的充电电池。\ce{ATP} 在细胞内的含量非常低,但它通过与 \ce{ADP}(腺苷二磷酸)之间的快速转化来满足巨大的能量需求:
- 水解(放能):ATP \rightarrow ADP + Pi + 能量。此过程为细胞的主动运输、肌肉收缩等提供动力。
- 再生(储能):ADP + Pi + 能量 \rightarrow ATP。能量主要来源于有机物的氧化分解(呼吸作用)或光合作用。
- 动态平衡:在一个典型的细胞中,\ce{ATP} 分子每 1-2 分钟就会被更新一次,人体每天水解并再合成的 \ce{ATP} 总量甚至相当于自身的体重。
能量利用的核心,能量偶联与磷酸化。细胞利用 \ce{ATP} 的核心策略是能量偶联(Energy Coupling),即利用 \ce{ATP} 水解这一放能反应来驱动原本无法自发进行的吸能反应:
- 磷酸化过程:\ce{ATP} 水解时,末端磷酸基团往往不是简单漂走,而是转移到另一个分子上,这一过程称为磷酸化。
- 改变构象:受体分子(如离子泵蛋白)在磷酸化后处于高能、不稳定的状态,从而发生构象改变,完成特定的生物学功能(如 Na⁺/K⁺ 泵的转运)。
广泛的应用实例:
- 主动运输:维持细胞内外的离子浓度梯度。
- 机械功:驱动肌肉纤维收缩、纤毛摆动或分子马达在细胞骨架上行走。
- 生物合成:连接单体以构建蛋白质、核酸等生物大分子。
- 特殊功能:如萤火虫发光(化学能转化为光能)、电鳐放电以及大脑思考。
细胞中绝大多数需要能量的生命活动都是由 \ce{ATP} 直接提供能量的,如大脑思考、电鳐发电和物质的主动运输都需要消耗 \ce{ATP}。\ce{ATP} 水解释放的能量是如何用于上述各种生命活动的呢?下图向你展示了 \ce{ATP} 是如何为主动运输供能的。
参与 \ce{Ca^2+} 主动运输的载体蛋白是一种能催化 \ce{ATP} 水解的酶。当膜内侧的 \ce{Ca^2+} 与其相应位点结合时,其酶活性就被激活了。
在载体蛋白这种酶的作用下,\ce{ATP} 分子的末端磷酸基团脱离下来与载体蛋白结合,这一过程伴随着能量的转移,这就是载体蛋白的磷酸化。
载体蛋白磷酸化导致其空间结构发生变化,使 \ce{Ca^2+} 的结合位点转向膜外侧,将 \ce{Ca^2+} 释放到膜外。
\ce{ATP} 水解释放的磷酸基团使蛋白质等分子磷酸化,这在细胞中是常见的。这些分子被磷酸化后,空间结构发生变化,活性也被改变,因而可以参与特定的化学反应。
课本:细胞的能量
细胞内的化学反应可以分成吸能反应和放能反应两大类。前者是需要吸收能量的,如蛋白质的合成等;后者是释放能量的,如葡萄糖的氧化分解等。许多吸能反应与 \ce{ATP} 水解的反应相联系,由 \ce{ATP} 水解提供能量;许多放能反应与 \ce{ATP} 的合成相联系,释放的能量储存在 \ce{ATP} 中,用来为吸能反应直接供能。也就是说,能量通过 \ce{ATP} 分子在吸能反应和放能反应之间流通。因此,可以形象地把 \ce{ATP} 比喻成细胞内流通的能量“货币”。正是由于细胞内具有 \ce{ATP} 这种能量“货币”,才能及时而持续地满足细胞各项生命活动对能量的需求。
我们现在来梳理一下:
直接能源物质:ATP。
主要能源物质:葡萄糖。
生物体内良好的储能物质:脂肪。
动物细胞内的储能物质:糖原。
植物细胞内的储能物质:淀粉。
最终能量来源:太阳(光)能。
对比实验:设置两个或两个以上的实验组,通过对结果的比较分析,来探究某种因素对实验对象的影响,这样的实验叫作对比实验,也叫相互对照实验。在本节课的探究活动中,需要设置有氧和无氧两种条件,探究酵母菌在不同氧气条件下细胞呼吸的方式,这两个实验组的结果都是事先未知的,通过对比可以看出氧气条件对细胞呼吸的影响。对比实验也是科学探究中常用的方法之一。
细胞供能的核心在于 \ce{ATP}(腺苷三磷酸),它被形象地称为细胞的“能量货币”。细胞通过将有机物(主要是葡萄糖)氧化分解,把释放的能量转移到 \ce{ATP} 中,从而驱动各项生命活动。根据氧气的参与情况,细胞供能主要分为以下几种方式:
有氧呼吸(需氧呼吸)是绝大多数生物获取能量的主要途径,能将葡萄糖彻底氧化分解,释放大量能量。1 mol 葡萄糖彻底氧化可释放 \pu{2870 kJ} 能量,其中约 34% 左右储存在 \ce{ATP} 中,其余以热能形式散失。
无氧呼吸(发酵):在缺氧条件下,细胞通过酶的催化将有机物不彻底分解,释放少量能量。
- 过程:第一阶段与有氧呼吸的糖酵解完全相同。第二阶段丙酮酸在不同酶的催化下转化为乳酸或酒精(并产生 \ce{CO2}),目的是再生糖酵解所需的 \ce{NAD+}。
- 类型:
- 乳酸发酵:见于乳酸菌、动物骨骼肌、马铃薯块茎等。
- 酒精发酵:见于酵母菌和大多数植物器官(如苹果、水稻根)。
- 能量产出:1 mol 葡萄糖分解成乳酸仅产生 2 分子 \ce{ATP}。
除了葡萄糖,细胞还可以代谢脂质和蛋白质:
- 脂肪:水解为甘油和脂肪酸。脂肪酸通过 \beta-氧化 产生乙酰辅酶 A 进入柠檬酸循环,产能量远高于糖类。
- 蛋白质:水解为氨基酸,脱去氨基后的碳链可进入有氧呼吸的不同环节进行氧化。
- 磷酸肌酸(肌肉特有):作为能量的“黄金储备”,能迅速将能量转移给 \ce{ADP} 来补充 \ce{ATP}。
| 能源物质 | 主要途径 | 场所 | 产能(每分子) |
|---|---|---|---|
| 葡萄糖 | 有氧呼吸 | 细胞质基质、线粒体 | 30 \sim 32 \ce{ATP} |
| 葡萄糖 | 无氧呼吸 | 细胞质基质 | 2 ATP |
| 脂肪酸 | \beta-氧化 | 线粒体 | ~108 \ce{ATP} (以软脂酸为例) |
这些机制共同构成了细胞的能量代谢网络。你是否想针对其中某一个具体过程(例如线粒体中的质子梯度是如何形成的)进行更深入的探讨?下面我们先综述,然后在下一节分别详细讲解。
对于绝大多数生物来说,有氧呼吸是细胞呼吸的主要形式,这一过程必须有氧的参与。有氧呼吸的主要场所是线粒体。线粒体具有内、外两层膜,内膜的某些部位向线粒体的内腔折叠形成嵴,嵴使内膜的表面积大大增加。嵴的周围充满了液态的基质。线粒体的内膜上和基质中含有许多种与有氧呼吸有关的酶。有氧呼吸最常利用的物质是葡萄糖,其化学反应式可以简写成:
\ce{C6H12O6 + 6H2O + 6O2 -> 6CO2 + 12H2O + 能量}
有氧呼吸的全过程十分复杂,可以概括地分为三个阶段,每个阶段的化学反应都有相应的酶催化。
第一个阶段是,1 分子的葡萄糖分解成 2 分子的丙酮酸,产生少量的 [\ce{H}],并且释放出少量的能量。这一阶段不需要氧的参与,是在细胞质基质中进行的。
第二个阶段是,丙酮酸和水彻底分解成二氧化碳和 [\ce{H}],并释放出少量的能量。这一阶段不需要氧直接参与,是在线粒体基质中进行的。
第三个阶段是,上述两个阶段产生的 [\ce{H}],经过一系列的化学反应,与氧结合形成水,同时释放出大量的能量。这一阶段需要氧的参与,是在线粒体内膜上进行的。
这里的 [\ce{H}] 是一种十分简化的表示方式。这一产生 [\ce{H}] 的过程主要是指氧化型辅酶 Ⅰ(\ce{NAD+})转化成还原型辅酶 Ⅰ(\ce{NADH})。
概括地说,有氧呼吸是指细胞在氧的参与下,通过多种酶的催化作用,把葡萄糖等有机物彻底氧化分解,产生二氧化碳和水,释放能量,生成大量 \ce{ATP} 的过程。同有机物在生物体外的燃烧相比,有氧呼吸具有不同的特点:有氧呼吸过程温和;有机物中的能量经过一系列的化学反应逐步释放;这些能量有相当一部分储存在 \ce{ATP} 中。
除酵母菌以外,还有许多种细菌(如乳酸菌)能够进行无氧呼吸。此外,马铃薯块茎、水稻根、苹果果实等植物器官的细胞以及动物骨骼肌的肌细胞等,除了能够进行有氧呼吸,在缺氧条件下也能进行无氧呼吸。一般地说,无氧呼吸最常利用的物质也是葡萄糖。
1 mol 葡萄糖在分解成乳酸以后,只释放出 \pu{196.65 kJ} 的能量,其中只有 \pu{61.08 kJ} 的能量储存在 \ce{ATP} 中,近 69% 的能量都以热能的形式散失了。人体肌细胞无氧呼吸产生的乳酸,能在肝脏中再次转化为葡萄糖。
无氧呼吸的全过程,可以概括地分为两个阶段,这两个阶段需要不同酶的催化,但都是在细胞质基质中进行的。第一个阶段与有氧呼吸的第一个阶段完全相同。第二个阶段是,丙酮酸在酶(与催化有氧呼吸的酶不同)的催化作用下,分解成酒精和二氧化碳,或者转化成乳酸。无论是分解成酒精和二氧化碳或者是转化成乳酸,无氧呼吸都只在第一阶段释放出少量的能量,生成少量 \ce{ATP}。葡萄糖分子中的大部分能量则存留在酒精或乳酸中。
无氧呼吸的化学反应式可以概括为以下两种:
\ce{C6H12O6 -> 2C3H6O3 (乳酸) + 少量能量}
\ce{C6H12O6 -> 2C2H5OH (酒精) + 2CO2 + 少量能量}
酵母菌、乳酸菌等微生物的无氧呼吸也叫作发酵。产生酒精的叫作酒精发酵,产生乳酸的叫作乳酸发酵。像这样,在没有氧气参与的情况下,葡萄糖等有机物经过不完全分解,释放少量能量的过程,就是无氧呼吸。
有氧呼吸和无氧呼吸都属于细胞呼吸。细胞呼吸是指有机物在细胞内经过一系列的氧化分解,生成二氧化碳或其他产物,释放能量并生成 ATP 的过程。所有生物的生存,都离不开细胞呼吸释放的能量。
细胞呼吸除了能为生物体提供能量,还是生物体代谢的枢纽。例如,在细胞呼吸过程中产生的中间产物,可转化为甘油、氨基酸等非糖物质;非糖物质代谢形成的某些产物与细胞呼吸中间产物相同,这些物质可以进一步形成葡萄糖。蛋白质、糖类和脂质的代谢,都可以通过细胞呼吸过程联系起来。
细胞呼吸的原理在生活和生产中得到了广泛的应用。生活中,馒头、面包、泡菜等许多传统食品的制作,现代发酵工业生产青霉素、味精等产品,都建立在对微生物细胞呼吸原理利用的基础上。在农业生产上,人们采取的很多措施也与调节呼吸作用的强度有关。例如,中耕松土、适时排水,就是通过改善氧气供应来促进作物根系的呼吸作用,以利于作物的生长;在储藏果实、蔬菜时,往往需要采取降低温度、降低氧气含量等措施减弱果蔬的呼吸作用,以减少有机物的消耗。
探究酵母菌细胞呼吸方式
探究酵母菌细胞呼吸方式的实验是高中生物学中的经典实验。通过该实验,我们可以认识到生物在不同供氧条件下获取能量的方式及其产物的差异。
实验材料:酵母菌的生物学特性。
生物类型:酵母菌是一种单细胞真菌,属于真核生物。
代谢类型:它是兼性厌氧菌。
- 在有氧条件下,它进行有氧呼吸,将葡萄糖彻底氧化分解为 \ce{CO2} 和水,并释放大量能量以供大量繁殖。
- 在无氧(缺氧)条件下,它进行酒精发酵,将葡萄糖分解为酒精和少量 \ce{CO2},释放少量能量。
实验原理与检测指标:实验通过检测呼吸产物(\ce{CO2} 和酒精)来判断呼吸方式。
二氧化碳(\ce{CO2})的检测
- 澄清石灰水:\ce{CO2} 可使澄清石灰水变浑浊。通过观察浑浊程度来比较 \ce{CO2} 的产生量。
- 溴麝香草酚蓝(BTB)水溶液:颜色变化为蓝 \rightarrow 绿 \rightarrow 黄。根据溶液变黄时间的长短可以检测 \ce{CO2} 的产生速度(变黄越快,说明 \ce{CO2} 产生越快、越多)。
酒精(乙醇)的检测
- 试剂:橙色的酸性重铬酸钾溶液(溶有重铬酸钾的浓硫酸溶液)。
- 现象:在酸性条件下,重铬酸钾与酒精反应变为灰绿色。
- 注意:葡萄糖也能与该试剂反应,因此检测前需延长培养时间以耗尽溶液中的葡萄糖。
实验设计:对比实验法。该实验属于对比实验(也叫相互对照实验)。
- 特点:设置两个或两个以上的实验组,通过对结果的比较分析,探究某种因素(本实验中为氧气)对实验对象的影响。
- 对比点:实验分为“有氧组”和“无氧组”,两组的结果在实验前都是未知的。
实验装置及关键操作:
有氧呼吸装置(甲组)
- 流程:空气 \rightarrow 10% NaOH 溶液 \rightarrow 酵母菌培养液 \rightarrow 澄清石灰水。
- NaOH 的作用:吸收空气中的 \ce{CO2},确保使石灰水变浑浊的 \ce{CO2} 全部来自酵母菌的有氧呼吸。
- 动力:连接橡皮球或气泵,保证 \ce{O2} 的充分供应。
无氧呼吸装置(乙组)
- 流程:酵母菌培养液(密封) \rightarrow 澄清石灰水。
- 关键操作:B 瓶(培养液瓶)应先封口放置一段时间,再连通盛有澄清石灰水的锥形瓶。
- 目的:让酵母菌消耗尽瓶中原有的 \ce{O2},确保检测到的 \ce{CO2} 是由无氧呼吸产生的。
| 呼吸方式 | \ce{CO2} 检测(石灰水/BTB) | 酒精检测(酸性重铬酸钾) |
|---|---|---|
| 有氧呼吸 | 浑浊程度高、变色快 | 不变色(橙色) |
| 无氧呼吸 | 浑浊程度低、变色慢 | 变灰绿色 |
实验结论:
- 酵母菌在有氧和无氧条件下都能进行细胞呼吸。
- 有氧条件下产生大量的 \ce{CO2} 和水;无氧条件下产生酒精和少量的 \ce{CO2}。
易错点拨(高考高频考点):
葡萄糖溶液煮沸:目的是杀菌和除去其中的 \ce{O2},防止杂菌干扰并利于创造无氧环境。
不能以“是否产生 \ce{CO2}”作为判断依据:因为两种呼吸方式都产生 \ce{CO2},必须依据产生量的多少或酒精的有无来判断。
酒精检测位置:检测液应取自培养液的滤液。若实验时间过短,葡萄糖未耗尽,会导致有氧组也可能出现灰绿色(伪阳性)。
糖异生、脂肪酸合成与 β-氧化
在生物化学代谢网络中,糖异生、脂肪酸合成与 \beta-氧化是维持机体能量稳态的核心路径。
一、糖异生:糖异生是指生物体利用非糖前体(如丙酮酸、乳酸、甘油和生糖氨基酸)合成葡萄糖的过程。在哺乳动物中,这一过程主要发生在肝脏中,少量发生在肾皮质。
- 生理意义:糖异生对于维持血糖水平至关重要,特别是为依赖葡萄糖供能的器官(如大脑和红细胞)提供持续的燃料。
- 核心步骤(绕过糖酵解的三个“旁路”):由于糖酵解中有三个步骤是不可逆的,糖异生必须通过特定的酶催化“旁路”反应来完成逆转:
- 旁路一:丙酮酸转变为磷酸烯醇式丙酮酸 (PEP)。这需要两步反应:首先在线粒体中由丙酮酸羧化酶(需生物素)将丙酮酸转化为草酰乙酸;随后草酰乙酸通过PEP羧激酶转化为PEP。
- 旁路二:果糖-1,6-二磷酸转变为果糖-6-磷酸。由果糖-1,6-二磷酸酶-1 (FBPase-1) 催化,该步骤是糖异生的关键调控点。
- 旁路三:葡糖-6-磷酸转变为游离葡萄糖。由葡糖-6-磷酸酶催化,该酶主要分布在肝和肾的内质网膜上。
- 能量消耗:糖异生是一个耗能过程。合成一分子葡萄糖需要消耗 4 分子 \ce{ATP} 和 2 分子 \ce{GTP}(总计 6 个高能磷酸键),以及 2 分子 \ce{NADH}。
- 代谢调节:糖异生与糖酵解受到正反交互调节,以避免“无效循环”。果糖-2,6-二磷酸是关键的别构信号分子,它激活糖酵解但抑制糖异生。此外,胰高血糖素会降低果糖-2,6-二磷酸的水平,从而刺激糖异生并抑制糖酵解。
二、脂肪酸合成:当机体能量供应充足时,多余的碳源会转化为脂肪酸并以三酰甘油的形式储存。
- 发生部位:主要发生在细胞的胞质溶胶中(植物在叶绿体基质中)。
- 前体准备:合成的主要原料是乙酰CoA。由于乙酰CoA产生于线粒体且无法直接跨膜,它需与草酰乙酸结合成柠檬酸穿梭进入胞质,再由柠檬酸裂解酶释放。
- 限速步骤:由乙酰CoA羧化酶 (ACC) 催化,将乙酰CoA转化为丙二酸单酰CoA。此反应需要生物素作为辅基并消耗ATP。
- 合成过程:由脂肪酸合酶 (FAS) 复合体催化。合成过程包含重复的四步循环:缩合、还原、脱水、再还原。
- 酰基载体蛋白 (ACP) 充当中间产物的载体。
- 还原反应的电子供体是 NADPH,主要由戊糖磷酸途径提供。
- 每次循环使碳链延长2个碳原子,直到合成 16碳的棕榈酸(软脂酸)。
- 调控机制:柠檬酸是ACC的别构激活剂,而棕榈酰CoA(最终产物)则是ACC的别构抑制剂。此外,胰高血糖素和肾上腺素会使ACC磷酸化而失活,抑制合成。
三、脂肪酸的 \beta-氧化:\beta-氧化是脂肪酸分解产生能量的主要途径,主要发生在线粒体基质中。
- 脂肪酸活化与转运:
- 活化:脂肪酸首先在胞质中由脂酰CoA合成酶催化,结合辅酶A形成脂酰CoA,此过程消耗ATP。
- 转运:长链脂酰CoA不能直接进入线粒体,需借助肉碱穿梭系统(肉碱脂酰转移酶I和II)通过线粒体内膜。丙二酸单酰CoA(脂肪酸合成的起始产物)可抑制肉碱脂酰转移酶I,防止合成与分解同时进行。
- 氧化过程:脂肪酸从羧基端开始,每轮循环切下两个碳原子(乙酰CoA)。每轮循环包含四步:脱氢、水合、再脱氢、硫解:
- 脱氢(一):由脂酰CoA脱氢酶催化,生成\ce{FADH2}。
- 水合:生成L-\beta-羟脂酰CoA。
- 脱氢(二):生成\ce{NADH}和\beta-酮脂酰CoA。
- 硫解:释放一分子乙酰CoA,并产生缩短了两个碳原子的脂酰CoA。
- 产能量:产生的乙酰CoA进入三羧酸循环 (TCA循环) 彻底氧化为CO₂,FADH₂和NADH进入呼吸链通过氧化磷酸化产生大量ATP。例如,一分子棕榈酸(C16)完全氧化约可产生106-108个ATP(视具体计算模型而定)。
- 特殊情况:
- 不饱和脂肪酸氧化需要额外的异构酶和还原酶。
- 奇数碳原子脂肪酸最后会产生丙酰CoA,需转化为琥珀酰CoA进入TCA循环。
- 酮体生成:在饥饿或糖尿病状态下,肝脏中过量的乙酰CoA会转化为酮体,为大脑等器官供能。
路径间的联系与调节总结:
- 互补与制约:糖类充足时,大量糖转化为脂肪酸(合成);当糖代谢障碍或供能不足时,脂肪酸动员并进行\beta-氧化。
- 关键调节点:丙二酸单酰CoA是脂肪酸合成的中间产物,它通过抑制肉碱转运系统来关闭\beta-氧化,确保代谢方向的单一性。
- 能量逻辑:等质量的脂肪比糖类氧化释放的能量多(约2倍多),是因为脂肪含氢量更高,含氧量极低,氧化时需氧量更大。
有氧呼吸的物质与产热
有氧呼吸是一个极其复杂的生物化学过程,通过一系列氧化还原反应,将葡萄糖中的化学能转化为生物能(\ce{ATP})。为了清晰梳理,我将有氧呼吸分为五个核心阶段:糖酵解、丙酮酸脱羧(链反应)、三羧酸循环、电子传递链(\ce{ETC})以及 \ce{ATP} 合酶(氧化磷酸化的最后部分)。
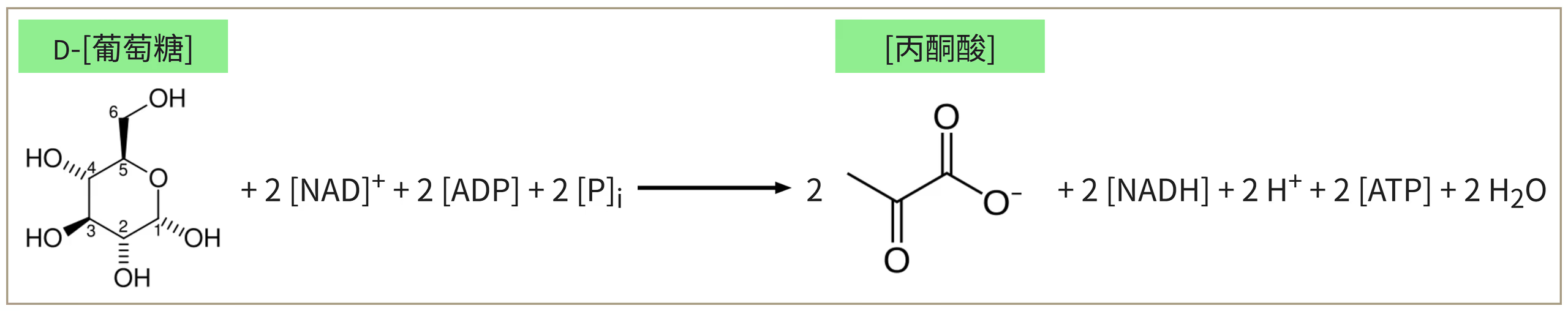
第一阶段:糖酵解。
细胞定位:细胞质基质。
核心化学反应:
\ce{C6H12O6 + 2NAD+ + 2ADP + 2Pi -> 2CH3COCOOH + 2NADH + 2H+ + 2ATP + 2H2O}
微观组分流向核算(每分子葡萄糖):
组分 消耗 生成 备注 碳链上的碳原子 6 6 一葡萄糖到两丙酮酸 \ce{CO2} 0 0 - \ce{H2O} 0 2 - 电子(\pu{e^-}) 0 4 转移到 2 个 \ce{NAD+} 上 质子(\ce{H+}) 0 4 2 个游离 \ce{H+},2 个结合在 \ce{NADH} 上 \ce{NADH} / \ce{NAD+} \ce{2NAD+} \ce{2NADH} 还原过程 \ce{ATP} / \ce{ADP} \ce{2ADP} \ce{4ATP} 净产物为 \ce{2 ATP}
第二阶段:丙酮酸氧化脱羧。
细胞定位:线粒体基质(丙酮酸通过线粒体内膜的丙酮酸载体 MPC 进入)。
核心化学反应:
\ce{2CH3COCOOH + 2NAD+ + 2CoA-SH -> 2CoA-S-COCH3 + 2CO2 + 2NADH + 2H+}
微观组分流向核算(每分子葡萄糖):
组分 消耗 生成 备注 碳链上的碳原子 6 4 两丙酮酸到两乙酰辅酶 \ce{A},2 个碳以 \ce{CO2} 释放 \ce{CO2} 0 2 脱羧反应,有氧呼吸释放 \ce{CO2} 的首个源头 \ce{H2O} 0 0 - 电子(\pu{e^-}) 0 4 转移到 2 个 \ce{NAD+} 上 质子(\ce{H+}) 0 4 2 个游离 \ce{H+},2 个结合在 \ce{NADH} 上 \ce{NADH} / \ce{NAD+} \ce{2NAD+} \ce{2NADH} - \ce{ATP} / \ce{ADP} - - -
第三阶段:三羧酸循环。
细胞定位:线粒体基质(唯有琥珀酸脱氢酶位于线粒体内膜上)
核心化学反应(两轮循环总和):
\ce{2CoA-S-COCH3 + 6NAD+ + 2FAD + 2GDP + 2Pi + 4H2O -> 4CO2 + 6NADH + 2FADH2 + 2GTP + 2CoA-SH + 6H+}
微观组分流向核算(每分子葡萄糖):
组分 消耗 生成 备注 碳链上的碳原子 4 0 两乙酰辅酶 \ce{A} 的碳全部以 \ce{CO2} 释放 \ce{CO2} 0 4 循环完全氧化 \ce{H2O} 4 0 宏观化学方程两轮共消耗 4 分子 电子(\pu{e^-}) 0 16 12 个给 \ce{NADH}、4 个给 \ce{FADH2} 质子(\ce{H+}) 0 16 6 个游离 \ce{H+},10 个结合在载体上(6在\ce{NADH},4在\ce{FADH2}) \ce{NADH} / \ce{NAD+} \ce{6NAD+} \ce{6NADH} - \ce{ATP} / \ce{ADP} - \ce{2ATP} 底物水平磷酸化
第四阶段:电子传递链。
细胞定位:线粒体内膜。
核心化学反应(多步串联总和):
\ce{10NADH + 10H+ + 2FADH2 + 6O2 -> 10NAD+ + 2FAD + 12H2O}
微观组分流向核算(每分子葡萄糖):
组分 消耗 生成 备注 \ce{O2} 6 0 最终电子受体 \ce{H2O} 0 12 还原产物 电子(\pu{e^-}) 24 0 20 个来自 \ce{NADH}、4 个来自 \ce{FADH2} 质子(\ce{H+}) 24 0 质子从基质被泵入膜间隙 \ce{NADH} / \ce{NAD+} \ce{10NADH} \ce{10NAD+} 汇集前三个阶段的全部产物 \ce{FADH2} / \ce{FAD} \ce{2FADH2} \ce{2FAD} 氧化过程
第五阶段:ATP 合酶与化学渗透。
细胞定位:线粒体内膜与大基质侧。
核心化学反应:
\ce{ADP} + \ce{Pi} + n\text{H}^+_{\text{out}} \longrightarrow \ce{ATP} + \ce{H2O} + n\text{H}^+_{\text{in}}
微观组分流向核算(每分子葡萄糖):
组分 消耗 (Consumption) 生成 (Generation) 备注 质子(\ce{H+}) 104\sim112 0 顺着浓度梯度跨膜回流 \ce{ATP} / \ce{ADP} 26\sim28[\ce{ADP + Pi}] 26\sim28[\ce{ATP}] 氧化磷酸化产生
关于水分子账单的深度校正说明:在化学层面上,\ce{ADP + Pi -> ATP + H2O} 的脱水缩合反应每生成一个 \ce{ATP} 就会产生一分子水(约 30 个 \ce{H2O})。然而,这些水分子在细胞后续发生 \ce{ATP} 水解做功时(\ce{ATP + H2O -> ADP + Pi})会被完全等量消耗。因此,在经典的生物大分子代谢账单中,为了聚焦于葡萄糖本身的碳氢骨架被氧化的过程,通常不计入高能磷酸键合成带来的水流,只核算碳骨架代谢与电子链形成的水。
基于前述不计高能键缩合水的原则,我们将全部小分子物质进行全面配平与对账:
| 微观具体物质/粒子 | 第一阶段:糖酵解 | 第二阶段:丙酮酸脱羧 | 第三阶段:三羧酸循环 | 第四阶段:氧化磷酸化 | 最终净收益/消耗 | 核心备注与生化机理 |
|---|---|---|---|---|---|---|
| 有机碳骨架 | 消耗 1 分子葡萄糖 \ce{C6},生成 2 分子丙酮酸 \ce{2C3} | 消耗 2 分子丙酮酸,生成 2 分子乙酰 \ce{2C2} | 消耗 2 分子乙酰 \ce{CoA} 完全拆解为无机碳 | 0 | 净消耗 1 分子葡萄糖 | 葡萄糖中的 6 个碳原子在第二、三阶段彻底被氧化拆解。 |
| 二氧化碳 | $0 | 生成 2 | 生成 4 | 0 | 净生成 6 | 2个来自丙酮酸脱氢酶复合体;4 个来自异柠檬酸与 \alpha-酮戊二酸的两次脱羧。 |
| 水分子 | 0 | 0 | 消耗 6 | 生成 12 | 净生成 6 | 无磷酸干扰下的纯账单:糖酵解无需水参与;为了把乙酰 \ce{CoA} 彻底变 \ce{CO2} 必须借 6 个水来补充氧原子;\ce{ETC} 生成 12 个水。 |
| 氧气 | 0 | 0 | 0 | 消耗 6 | 净消耗 6 | 仅在第四阶段作为呼吸链末端的最终电子与质子受体。 |
| 高能电子 | 剥离 4 个 | 剥离 4 个 | 剥离 16 个 | 消耗 24 个 | 动态平衡 | 共 24 个高能电子被脱氢酶剥离,由辅酶携带后悉数注入内膜呼吸链,最终交给氧气。 |
| 质子 | 剥离 4 个 | 剥离 4 个 | 剥离 16 个 | 化学消耗 24 个,物理跨膜回流约 104\sim 112 个 | 动态平衡 | 化学反应:24 个质子最终与 \ce{O2}、\ce{e^-} 结合生成水。物理做功:泵出的质子经 \ce{ATP} 合酶顺浓度梯度回流做功。 |
| \ce{NADH} | 生成 2 | 生成 2 | 生成 6 | 消耗 10 | 动态平衡 | 细胞质中产生的 2 个 \ce{NADH} 需经穿梭系统(苹果酸-天冬氨酸或甘油-3-磷酸)进入线粒体电子传递链。 |
| \ce{FADH2} | 0 | 0 | 生成 2 | 消耗 2 | 动态平衡 | 结合在线粒体内膜琥珀酸脱氢酶(复合体 II)上直接传递电子。 |
| \ce{ATP} / \ce{GTP} | 净收益 2 | 0 | 生成 2 \ce{GTP} | 生成 26 \sim 28 | 净生成 30 \sim 32 | 前三个阶段均为底物水平磷酸化;末端为基于质子动力的氧化磷酸化。 |
能量转换效率与现代生化模型的定量修正:为什么最终生成的 \ce{ATP} 是一个范围值(30 或 32)?这主要取决于细胞质中产生的 2 分子糖酵解 \ce{NADH} 跨过线粒体内膜时的穿梭机制:
苹果酸-天冬氨酸穿梭:主要存在于哺乳动物的心肌、肝脏和肾脏中。此穿梭不损失能量,细胞质中的 \ce{NADH} 相当于直接转运为线粒体内部的 \ce{NADH}。其电子进入复合体 \ce{I},因此可正常泵出 10 个 \ce{H+},每分子产生约 2.5 个 \ce{ATP}。此时总产量为 \ce{32ATP}。
甘油-3-磷酸穿梭:主要存在于骨骼肌和脑组织中。其电子跨膜后交给了线粒体内膜上的甘油-3-磷酸脱氢酶,直接将电子传递给辅酶 \ce{Q}(相当于绕过了复合体 \ce{I},以 \ce{FADH2} 形式入场)。因此每分子只能泵出 6 个 \ce{H+},产生约 1.5 个 \ce{ATP}。此时总产量降为 \ce{30 ATP}。
旧教材中的 36 或 \ce{38 ATP} 是如何得来的?:
在早期的生物化学中,科学家估算的 \text{P}/\text{O} 比值(即每消耗一个氧原子所磷酸化的 \ce{ADP} 数量)为整数。当时认为 1 摩尔 \ce{NADH} 恒定生成 3 摩尔 \ce{ATP},1 摩尔 \ce{FADH2} 恒定生成 2 摩尔 \ce{ATP}。计算公式为 (2+2) + 10 \times 3 + 2 \times 2 = \pu{38 ATP},或因穿梭损耗变为 36。
自 20 世纪 90 年代以来,结合 \ce{ATP} 合酶 c 环结构的精细解析和质子动力势的严密测定,公认的非整数实际 \text{P}/\text{O} 比修正为:\ce{NADH} \approx \pu{2.5 ATP},\ce{FADH2} \approx \pu{1.5 ATP}。由此推导出的 30 或 \ce{32 ATP} 更加符合真实的细胞生理状况。
在标准状态下,1 摩尔葡萄糖彻底氧化释放的吉布斯自由能约为 \pu{-2870 kJ}。而在细胞内生理条件下,合成 1 摩尔 \ce{ATP} 所需的标准自由能约为 \pu{30.5 kJ/mol}。但在活细胞真实的生理状态下(受 \ce{ATP}/\ce{ADP} 极高浓度比值影响),实际合成 1 摩尔 \ce{ATP} 需要克服的自由能约为 46 \sim 52\text{ kJ/mol}。
- 理论标准效率(常用于传统教材算账):\eta = \dfrac{32 \times 30.5\text{ kJ}}{2870\text{ kJ}} \times 100\% \approx \mathbf{34\%}
- 实际生理效率:\eta \approx \dfrac{32 \times 50\text{ kJ}}{2870\text{ kJ}} \times 100\% \approx \mathbf{55\%} (表明细胞其实是一台效率极高的能量转化机器)。
能量去向归宿:
- 约 34\% 的热力学自由能被成功俘获并锁定在高能磷酸键中,供细胞完成各项生命活动(如主动运输、肌肉收缩、大分子合成等)。
- 其余约 66\% 的自由能以热能的形式散失。这一部分热能用于维持恒温动物的核心体温。保持这种热散失也符合热力学第二定律,确保了整个细胞内宏观有氧呼吸反应流的自发性(\Delta G < 0)。
有氧呼吸是生物体获取能量的核心过程,其调节方式不仅涉及细胞内的代谢平衡,还与整体的体温维持(产热)密切相关。有氧呼吸是指细胞在氧的参与下,通过多种酶的催化作用,把葡萄糖等有机物彻底氧化分解,产生二氧化碳和水,释放能量并生成大量 \ce{ATP} 的过程。
- 能量分配:有机物分解释放的能量,大部分(约 60\%\sim69\%)以热能的形式散失,少部分储存在 \ce{ATP} 中供生命活动利用。
- 意义:呼吸作用释放的热能用于维持体温(恒温动物),而 \ce{ATP} 则直接驱动肌肉收缩、物质运输等各项生命活动。
哺乳动物有氧呼吸与产热的调节:在人等哺乳动物中,呼吸速率和产热受细胞代谢需求、神经系统和内分泌系统的共同精细调节。在细胞水平的自我调节(受体控制):
\ce{ADP}/\ce{ATP} 比率:线粒体中呼吸的速率受 \ce{ADP} 浓度的严格限制。当能量消耗增加(如肌肉收缩),\ce{ATP} 水解产生更多 \ce{ADP},\ce{ADP} 进入线粒体作为磷酸化底物,直接刺激呼吸速率增加。这种根据 \ce{ADP} 需求来调节呼吸的机制被称为受体控制。
关键酶调节(PFK-1):磷酸果糖激酶 1(PFK-1)是有氧呼吸第一阶段的关键酶。细胞内 \ce{ATP}/\ce{AMP} 的浓度比变化会调节其活性:当 \ce{ATP} 充足时会抑制该酶,减缓呼吸;当 \ce{ATP} 减少、\ce{AMP} 增多时(如运动时),会激活该酶,加快细胞呼吸速率以产生更多能量。
植物通过环境因素调节呼吸,并演化出特殊的生热机制。环境因素的影响:
温度:主要影响酶活性。在最适温度范围内,呼吸速率随温度升高而增大;超过最适温度,酶活性降低甚至变性,呼吸速率下降。
氧气浓度:\ce{O2} 是有氧呼吸的必需原料,同时抑制无氧呼吸。低氧条件下有氧呼吸受抑,但在保存果蔬时,适当低氧可减少有机物消耗。
\ce{CO2} 浓度:\ce{CO2} 是呼吸终产物,积累过多会产生反馈抑制效应,抑制呼吸作用。
特殊产热机制(交替氧化酶途径):
生热植物(如臭菘):某些植物(如天南星科的臭菘、海芋)在开花期肉穗花序会显著发热,温度可比环境高出 20\sim\pu{40^oC}。
机制:这些植物线粒体具有交替氧化酶(AOX)。电子在传递链中绕过产生 \ce{ATP} 的质子泵(复合体 III 和 IV),直接从泛醌传递给氧。电子传递释放的所有能量都转化为热能,用于挥发气味吸引昆虫授粉,或在早春融化冰雪。
关键代谢效应:巴斯德效应与瓦堡效应。
巴斯德效应:在既能发酵又能呼吸的微生物(如酵母菌)中,**氧气抑制糖酵解(发酵)**的现象。
在有氧条件下,细胞通过有氧呼吸产生 \ce{ATP} 的效率远高于无氧呼吸(消耗等量葡萄糖,产生的 \ce{ATP} 相差约 15\sim18 倍)。高效的 \ce{ATP} 产生会导致细胞内 \ce{ATP}/\ce{AMP} 比率升高,进而抑制糖酵解的关键酶(PFK-1),使得葡萄糖消耗速率大幅下降。
瓦堡效应:癌细胞即使在氧气充足的条件下,也主要依赖无氧呼吸(产生乳酸)来产生 $\ce{ATP} 的现象。
- 快速供能:虽然无氧呼吸产能效率低,但其产生的中间产物可转化为氨基酸、核苷酸等,为癌细胞的快速分裂提供物质原料。
- 调节异常:癌细胞通过大量吸收葡萄糖(吸收速率比正常细胞快得多)来弥补无氧呼吸产能效率的不足。这通常涉及细胞内信号通路的改变,使其代谢模式从高效的有氧氧化转向快速的“倍增模式”。
代谢途径
代谢途径,在生物化学中,是生物体将一种物质通过一系列酶的顺序作用(酶促反应)而转化成另一种代谢物的反应序列,包括合成或分解的过程。代谢途径由多个连续的代谢反应相互连接起来,进而完成物质的合成或分解,属于化学反应的一种。细胞内不同代谢途径组成了代谢网络。底物是否进入代谢途径,要视乎细胞的需要,即合成代谢物及分解代谢物浓度的独特组合(流量控制反应的动力)。代谢途径包括主要的代学反应(一般都是需要酶的)令生物保持它的内环境稳态。
代谢途径一般都有以下特征:它们包括多个步骤,就像瀑布一样。首个步骤多数不能倒逆。其他步骤并不一定不能倒逆。大部分情况下,这些步骤可以视乎细胞的实际需要而倒逆。糖酵解是极好的例子:当葡萄糖进入细胞后,它立即会被三磷酸腺苷(\ce{ATP})磷酸化,在首个不能倒逆的步骤成为葡萄糖-6-磷酸。这是要避免葡萄糖离开细胞。在过多脂类或蛋白质时,糖酵解会倒逆进行糖异生,以产生葡萄糖六磷酸盐作为糖原或淀粉储存。它们都是有调节的,一般是以反馈方式,或是以循环形式再次进行反应(如三羧酸循环)。真核生物的合成代谢及分解代谢途径,大都局限于细胞中的特定区域(细胞区室),或是以使用不同的酶或辅助因子来分开。
呼吸作用是生物体细胞把有机物氧化分解并转化能量的化学过程,该过程也称为释放作用。无论是否为自养,细胞内完成生命活动所需要的能量都是来自呼吸作用。在真核细胞中,线粒体是与呼吸作用密切相关的细胞器,呼吸作用的几个关键性步骤都在其中进行。呼吸作用是一种酶促氧化反应。虽名为氧化反应,不论有否氧气参与,都可称作呼吸作用(这是因为在化学上,有电子转移的反应过程,皆可称为氧化)。有氧气参与时的呼吸作用,称之为有氧呼吸;没氧气参与的反应,则称为无氧呼吸。
呼吸作用的目的,是透过释放食物里之能量,以制造三磷酸腺苷,即细胞最主要的直接能量供应者。呼吸作用与氢氧的燃烧之间最大分别是:呼吸作用透过一连串的反应步骤产生能量(一部分储存于 \ce{ATP} 中),而燃烧则是将能量一次性的释放。在呼吸作用中,三大营养物质:碳水化合物、蛋白质和脂质的基本组成单位──葡萄糖、氨基酸和脂肪酸,被分解成更小的分子,并透过数个步骤,将能量转移到 \ce{NADH} 中。最后经过一系列的电子传递链,\ce{NADH} 与氧气共同作用生成水,而原本贮存在其中的能量,则转移到 \ce{ATP} 分子上,以供生命活动使用。
包扎伤口时,需要使用透气的消毒纱布或 OK 绷等敷料。以避免厌氧菌的繁殖,如破伤风芽孢杆菌。这种病菌只能进行无氧呼吸,皮肤破损较深或被锈钉扎伤病菌就容易大量繁殖。利用麦芽糖,葡萄糖,粮食和酵母菌以及发酵罐等,在控制通气的情况下可以生产各种酒;利用淀粉醋酸杆菌或谷氨酸棒状杆菌以及发酵罐控制通气的情况下,可以生产食醋或味精。花盆里的土壤板结后空气不足会影响根系生长,需要及时松土透气,有利于植物更好吸收氧气。稻田定期排水,避免水稻幼根因缺氧而变黑腐烂。提倡慢跑等有氧运动的原因之一是不致因剧烈运动而导致氧的不足,二是肌细胞无氧呼吸产生大量乳酸。乳酸的大量积累会使肌肉酸胀乏力。
最基础的代谢途径:糖酵解
糖酵解(Glycolysis)是生物体获取能量的基础代谢途径,具有极高的保守性,几乎存在于地球上所有的细胞中,证明了它起源于生命史的早期。根据维基百科,糖酵解分为十个阶段,为了更符合高考情景,这里简化为大约 7 个步骤。
糖酵解发生在细胞的细胞质基质(胞质溶胶)中,无论是原核生物还是真核生物均是如此。该过程不需要氧气,属于厌氧代谢,是细胞在缺氧环境下唯一的能量来源(如成熟的哺乳动物红细胞)。
准备阶段:这些步骤消耗能量以将葡萄糖转变为两个丙糖磷酸,即甘油醛-3-磷酸和磷酸二羟丙酮。此阶段共投入两分子 \ce{ATP}。
在己糖激酶(一种转移酶)的催化下,消耗一分子 \ce{ATP},将一分子葡萄糖(\ce{C6})转化为一分子葡萄糖-6-磷酸(\ce{C6-P})。辅因子:镁离子。
而肝脏中,存在于一种特殊的己糖激酶名叫葡萄糖激酶,这种酶只对葡萄糖有专一性,并不会与所有己糖发生作用,而且此酶并不受葡萄糖-6-磷酸的抑制作用。它只在葡萄糖浓度很高时才作用,由于肝脏是糖原的重要生成器官,因此当血糖浓度增高时,肝脏中的葡萄糖激酶就运作,生成了葡萄糖-6-磷酸,肝脏以此合成肝糖原。
此酶的合成受到胰岛素的诱导,因此,体内缺乏胰岛素的糖尿病患者的肝脏中此酶的合成速率低,影响肝脏中葡萄糖转变为葡萄糖-6-磷酸的速率,故分解葡萄糖以及合成肝糖原的过程受阻。
在磷酸葡萄糖异构酶(一种异构酶)的催化下,一分子葡萄糖-6-磷酸(\ce{C6-P})异构化,转化为对称的一分子果糖-6-磷酸(\ce{C6-P}),为裂解做准备。
在磷酸果糖激酶(\ce{PFK},一种转移酶)的催化下,消耗一分子 \ce{ATP},将一分子果糖-6-磷酸(\ce{C6-P})转化为一分子果糖-1,6-二磷酸(\ce{P-C6-P})。辅因子:镁离子。
在细胞内磷酸果糖激酶反应为不可逆的,磷酸果糖激酶属于一种调节性酶,且为肌肉糖解作用中主要的调节性酶,受到 \ce{ATP} 与 \ce{AMP} 数量比的影响:当 \ce{ATP} 过多时,\ce{ATP} 结合到酶的调控部位,酶的构象发生变化而受抑制;\ce{AMP} 会解除此酶的抑制,使其恢复高效状态。
氢离子浓度对此酶也有抑制作用:当血液中的乳酸较多时,即氢离子浓度增高,这样就抑制了酶,使得糖酵解效率下降,阻断了糖酵解下游生成乳酸的途径,因此这种调控有着重要的生理意义。
前一步反应使得分子失稳,在果糖二磷酸醛缩酶(一种裂解酶)的催化下,己糖环——一分子果糖-1,6-二磷酸(\ce{P-C6-P})被醛缩酶分成两个丙糖,磷酸丙糖异构酶(一种异构酶)迅速异构化二羟丙酮磷酸,最终产生两分子甘油醛-3-磷酸(\ce{P-C3})。
放能阶段:糖解作用的第二阶段为放能阶段,此阶段的目的在于产生高能分子 \ce{ATP} 和 \ce{NADH}。因为一个葡萄糖在准备阶段时已经变成两个丙糖,所以在放能阶段中每个反应会发生两次。最后产生 2 个 \ce{NADH} 和 4 个 \ce{ATP},使得单一葡萄糖在经过整个糖解作用后净得 2 个 \ce{NADH} 和 2 个 \ce{ATP}。\ce{ATP} 会用于其他需能反应,而 \ce{NADH} 则会进入呼吸链或作为还原剂参与细胞内其他还原加氢反应。
在甘油醛-3-磷酸脱氢酶(\ce{GAPDH})的催化下,两分子甘油醛-3-磷酸(\ce{P-C3})被氧化并各添加一分子无机磷酸,产生两分子甘油酸-1,3-二磷酸(\ce{P-C3-P})。同时,脱下的两分子氢用于还原氢载体两分子 \ce{NAD+},形成两分子 \ce{NADH}。
在磷酸甘油酸激酶(\ce{PGK},一种转移酶)的催化下,两分子甘油酸-1,3-二磷酸(\ce{P-C3-P})分别脱去各一个磷酸基团,转移至 \ce{ADP},生成两分子 \ce{ATP} 和两分子甘油酸-3-磷酸(\ce{P-C3})。
在这一步,糖酵解过程达到了能量收支平衡:2 分子 \ce{ATP} 在先前的反应中被消耗,而在这步反应中有两分子 \ce{ATP} 被合成。这步反应作为两步底物水平磷酸化中的一步,以 \ce{ADP} 作为底物,所以当细胞 \ce{ATP} 水平较高时,该步反应被抑制;因此该步反应也是糖酵解过程中重要的控速步骤之一。
分别通过磷酸甘油酸变位酶(\ce{PGM},一种变为酶)生成甘油酸-2-磷酸,烯醇化酶(\ce{ENO},一种裂解酶)脱去一分子水生成磷酸烯醇式丙酮酸,丙酮酸激酶(\ce{PK})最终生成丙酮酸并将一分子 \ce{ADP} 转化为一分子 \ce{ATP}。
第二阶段较为复杂,可以简单记为:每一分子 \ce{P-C3} 被氧化,脱下两个质子和两个电子,质子和电子转移给一分子 \ce{NAD+} 生成一分子 \ce{NADH},同时,高能中间产物将磷酸基团(以及额外的两分子 \ce{Pi})转移给两分子 \ce{ADP} 生成两分子 \ce{ATP}。在此阶段,一共有两分子一磷酸三碳糖参与反应,共消耗两分子 \ce{NAD+}产生两分子 \ce{NADH},消耗两分子 \ce{ADP} 和两分子 \ce{Pi} 产生两分子 \ce{ATP}。
能量统计与代谢调节:
于能量而言:投入两分子 \ce{ATP},产出四分子 \ce{ATP} 和两分子 \ce{NADH},净收益为两分子 \ce{ATP} 和两分子 \ce{NADH}。
于物质而言:此阶段不消耗水、产生两分子水,消耗两分子 \ce{Pi} 将两分子 \ce{ADP} 转化为两分子 \ce{ATP}。
于细胞而言:此阶段在细胞质基质中进行,无须细胞器参与(哺乳动物成熟的红细胞主要依赖糖酵解功能),在有氧、无氧条件下均可以进行。
精密调节:细胞根据能量状态实时调控,2024 年安徽卷第 3 题据此出题。当 \ce{ATP} 充足或柠檬酸积累时,会抑制 PFK 的活性,让糖酵解“减速”;当 \ce{ADP} 或 \ce{AMP} 增多时,则激活 PFK,加速产能。
糖酵解的终点是丙酮酸,它的命运由氧气决定:
- 有氧条件下:丙酮酸进入线粒体基质,氧化脱羧生成乙酰辅酶 A,进而参与三羧酸循环,产生大量的 \ce{ATP}。
- 无氧条件下(发酵):为了让糖酵解不因为缺少 \ce{NAD+} 而中断,细胞必须将 \ce{NADH} 重新氧化。丙酮酸在不同酶的作用下转化为乳酸(动物、乳酸菌)或乙醇和 \ce{CO2}(酵母、多数植物)。
糖酵解不仅是产能过程,它产生的许多中间产物还为合成氨基酸、脂质和核苷酸提供了重要的碳骨架。
丙酮酸的去向:丙酮酸脱羧和无氧呼吸
丙酮酸脱羧是连接糖酵解与三羧酸循环的关键“桥梁反应”,在代谢调节中占据核心地位。在真核细胞中,这一过程发生在线粒体基质中。催化这一反应的是一个巨大的丙酮酸脱氢酶复合体(\ce{PDC}),它由三种酶(\ce{E1}、\ce{E2}、\ce{E3})组成,其体积甚至比核糖体还要大。该复合体在氧化脱羧过程中需要多种维生素衍生物作为辅酶,包括来自维生素 \ce{B1} 的 \ce{TPP} 和来自维生素 \ce{B5} 的辅酶 \ce{A}(\ce{CoA})。
在 \ce{PDC} 的精密配合下,一分子丙酮酸(\ce{C3})最终转化为 1 分子乙酰辅酶 \ce{A}(\ce{C2}):我们在化学中学过脱羧反应,与丙酮酸 \ce{CH3C(O)COOH} 脱羧形成 \ce{CH3COH} 乙醛类似,丙酮酸与 \ce{TPP} 结合,脱去 \ce{CO2} 形成羟乙基,随后羟乙基被氧化为乙酰基,脱下的电子通过还原一分子 \ce{NAD+} 产生一分子 \ce{NADH},为后续的氧化磷酸化储备高能电子。活化的乙酰基被转移到辅酶 \ce{A}(\ce{CoA-SH})上,形成乙酰辅酶 \ce{A}(\ce{CoA-S-COCH3})。
这一反应是细胞代谢的一个决定性时刻:
- 不可逆性:在动物体内,丙酮酸一旦转化为乙酰辅酶 \ce{A},就无法再通过糖异生变回葡萄糖。
- 枢纽作用:它不仅处理糖类代谢产物,脂肪酸的 \beta-氧化也会产生乙酰辅酶 \ce{A} 进入此路径,使三羧酸循环成为三大营养物质氧化的共同通路。
- 去向选择:当细胞能量(\ce{ATP})短缺时,乙酰辅酶 \ce{A} 进入循环彻底氧化;当能量充足时,它可转变为柠檬酸并回到细胞质中用于合成脂肪。
另一种路径:简单脱羧。在缺氧环境下的酒精发酵中,丙酮酸不进行氧化脱羧,而是进行无氧发酵。
无氧呼吸是细胞在无氧条件下,通过酶的催化作用将有机物不彻底氧化分解,释放少量能量并产生 \ce{ATP} 的过程。这一过程完全发生在细胞质基质中,可以分为糖酵解和丙酮酸还原。
由于细胞所含酶的种类不同,丙酮酸在第二阶段会有不同的去向:
酒精发酵(Alcoholic Fermentation):
- 过程:丙酮酸首先在丙酮酸脱羧酶的作用下脱去 \ce{CO2} 生成乙醛,乙醛再被 \ce{NADH} 还原为乙醇。
- 反应式:\ce{C6H12O6 -> 2C2H5OH + 2CO2 + 少量能量}。
- 典型生物:酵母菌以及大多数植物在缺氧时(如淹水的根系或苹果果实)。
乳酸发酵(Lactic Acid Fermentation):
- 过程:丙酮酸在乳酸脱氢酶的催化下,直接被 \ce{NADH} 还原为乳酸,此过程不产生 \ce{CO2}。
- 反应式:\ce{C6H12O6 -> 2C3H6O3 + 少量能量}。
- 典型生物:乳酸菌、高等动物剧烈运动时的骨骼肌,以及少数植物器官(如马铃薯块茎、玉米胚、甜菜块根)。
生理意义与核心机制:
- \ce{NAD+} 再生:发酵的关键不在于产生更多能量,而在于将糖酵解产生的 \ce{NADH} 重新氧化为 \ce{NAD+},从而保证糖酵解能够持续进行并提供最基本的 \ce{ATP}。
- 能量利用率低:葡萄糖中的大部分能量仍储存在酒精或乳酸中,未被完全释放。
无氧呼吸的产物(酒精和乳酸)可以进一步分解供能,或者在一定条件下转化为其他能源物质再利用。无氧呼吸被称为“不彻底的氧化分解”,其主要原因就是葡萄糖分子中的大部分能量仍然储存在产物酒精或乳酸中,只有少部分能量被释放出来用于合成 \ce{ATP}。
乳酸的进一步分解与再利用:在哺乳动物(包括人)中,乳酸的代谢主要通过血液循环与肝脏的协同作用完成。
- 转化为葡萄糖(\ce{Cori} 循环):剧烈运动时,骨骼肌无氧呼吸产生的乳酸会进入血液,并被运送到肝脏。在肝脏中,乳酸可以通过糖异生作用转化回丙酮酸,进而重新合成葡萄糖或糖原。这一过程不仅消耗了积累的乳酸,维持了内环境 \pH 的稳定,还实现了能量的回收利用。
- 进入有氧氧化:如果氧气供应变得充足,乳酸可以在乳酸脱氢酶(\ce{LDH})的催化下转化回丙酮酸,然后进入线粒体进行三羧酸循环,被彻底氧化分解为二氧化碳和水,并产生大量的 \ce{ATP}。
酒精(乙醇)的进一步分解与利用:酒精也是一种富含能量的分子(它是优良的燃料),可以被生物体进一步氧化。
- 肝脏中的氧化降解:在人类等脊椎动物体内,从小肠吸收或代谢产生的酒精主要在肝脏中处理。通过乙醇脱氢酶(\ce{ADH})的作用,酒精首先被氧化为乙醛,随后进一步转化为乙酸(醋酸),最终生成乙酰辅酶 \ce{A}。乙酰辅酶 \ce{A} 可以进入三羧酸循环彻底氧化供能,或者转化为脂肪储存。
- 微生物的二次利用:在一些微生物(如酵母菌)中,虽然酒精是其发酵的产物,但当环境中的葡萄糖耗尽且氧气充足时,它们也可以“反过来”利用酒精作为碳源和能源进行有氧呼吸。
由于无氧呼吸产物中仍含有大量未释放的能量,因此这些产物在有氧条件下具有很高的再利用价值。但需要注意的是,酒精对细胞具有毒害作用,如果积累过多(如植物烂根或酵母菌酒精中毒),会导致细胞受损或死亡。
有氧呼吸的核心:三羧酸循环
三羧酸循环(TCA 循环)是有氧呼吸的核心,它不仅是能量产生的“主发动机”,更是细胞内各种物质代谢的枢纽。“三羧酸循环”之所以得名,是因为该循环中最重要的初始产物——柠檬酸是一种含有三个羧基(\ce{-COOH})的有机酸。
注意:在真核细胞中,这一循环完全发生在线粒体基质中,虽然三羧酸循环本身不消耗氧气,但是在无氧环境下不会发生。
在循环开始前,糖酵解产生的丙酮酸必须通过线粒体膜进入基质,丙酮酸脱羧过程,最终转化为乙酰辅酶 \ce{A}(\ce{CH3CO-CoA})。乙酰辅酶 \ce{A} 就像一张“入场券”,将碳骨架送入循环。
为了展现出碳的变化,在三羧酸循环中经常用 \ce{C_x} 表示碳链上碳原子数为 x 的某种有机物,例如 \ce{C1} 是 \ce{CO2},\ce{C2} 是乙酰辅酶 \ce{A} 等。
三羧酸循环通常被视为有八个特定酶促反应组成,且相当复杂,为了简便,用碳链上碳原子数划分为三个大阶段:
柠檬酸合成酶催化乙酰辅酶 \ce{A}(\ce{CoA-S-COCH3},记为 \ce{C2})与草酰乙酸(\ce{C4})发生缩合反应和硫酯水解,生成柠檬酸(\ce{C6})。
\ce{CoA-S-COCH3 + C4 -> CoA-SH + C6}
该反应高度放能(硫酯水解释放能量),也因此该步骤在标准条件下不可逆。此外,酶产生的柠檬酸能够竞争性抑制酶活性,尽管该反应非常有利(因为是放能的),但柠檬酸合酶可以受到严格调节。例如,柠檬酸盐本身充当柠檬酸合成酶的抑制剂,而草酰乙酸与其结合会增加其对乙酰辅酶A的亲和力。这方面具有相当大的生物学重要性,因为它可以完全调节整个克雷布斯循环,使酶成为整个循环的“起搏器”。糖酵解中的磷酸果糖激酶会被柠檬酸盐抑制,同时它会激活乙酰辅酶 \ce{A} 羧化酶以进行脂肪酸合成。这一点说明了我们代谢周期的相互关联性。
在若干酶的催化下,柠檬酸(\ce{C6})经过一系列反应(包含一次脱羧)生成 \alpha-酮戊二酸(\ce{C5})、再经过一系列反应(包含一次脱羧)生成琥珀酸(\ce{C4})。在 \alpha-酮戊二酸 \ce{C5} 前后还有辅酶 \ce{A} 的参与。
\ce{C6 + 2NAD+ + GDP + Pi -> C5 + CO2 + 2NADH + 2H+ + GTP}
\ce{C6 + 2NAD+ + ADP + Pi -> C5 + CO2 + 2NADH + 2H+ + ATP}
这一部分整个过程,共释放两分子 \ce{CO2},合成两分子 \ce{NADH} 和一分子 \ce{ATP} 或 \ce{GTP}(该酶有两个版本,分别生成 \ce{ATP} 和 \ce{GTP};不同生物,或同一生物的不同组织会依对 \ce{ATP} 和 \ce{GTP} 需求的差异在该步使用相应版本的酶)。
循环的最后部分是四碳分子的重新排列,直到草酰乙酸。为实现这一点,琥珀酸上存在的亚甲基桥必须转化为羰基,这种转化通过三个步骤发生:第一次氧化、水合和二次氧化。这三个步骤,除了再生草酰乙酸之外,还可以通过形成 \ce{FADH2} 和 \ce{NADH} 来提取更多能量。
第一个氧化反应是琥珀酸在琥珀酸脱氢酶的催化作用下,转化为延胡索酸。该过程中有一分子 \ce{FAD} 还原为 \ce{FADH2}。
延胡索酸经延胡索酸酶催化,催化一分子水分子加成到反式分子上,转化为苹果酸。
苹果酸在苹果酸脱氢酶的催化作用下,转化为草酰乙酸。该过程生成一分子 \ce{NADH}。至此,柠檬酸循环完成。生成的草酰乙酸可再次进入柠檬酸循环。
琥珀酸脱氢酶复合物是循环中唯一以 \ce{FAD} 而不是 \ce{NAD+} 作为氢受体的酶,\ce{FAD} 通过组氨酸残基与酶共价连接。该酶使用 \ce{FAD},因为与反应相关的能量不足以还原 \ce{NAD+}。
与最后一个反应相关的吉布斯自由能显然是正的(与循环中的其他反应不同)。该酶的活性是由柠檬酸合酶消耗草酰乙酸和电子传递链消耗 \ce{NADH} 驱动。该反应平衡倾向于苹果酸一侧,但因草酰乙酸不断消耗,反应得以持续进行。基于上述理由,该反应也是柠檬酸循环的一个重要调控位点。
高等动物的柠檬酸循环可用下式表示:
\ce{CoA-S-COCH3 + 3NAD+ + FAD + GDP + Pi + 2H2O -> CoA-SH + 3NADH + 3H+ + FADH2 + GTP + 2CO2}
\ce{CoA-S-COCH3 + 3NAD+ + FAD + ADP + Pi + 2H2O -> CoA-SH + 3NADH + 3H+ + FADH2 + ATP + 2CO2}
能量统计与代谢产出:每消耗一分子乙酰辅酶 \ce{A}(对应半个葡萄糖),完整的三羧酸循环进行一轮,草酰乙酸的分子数目不会发生变化,因此,从净反应的意义上讲,可以认为草酰乙酸起了催化剂的作用。循环的净产出为:
- 三分子 \ce{NADH} 和一分子 \ce{FADH2}(高中通常直接记作 [\ce{H}] 为一分子 \ce{NADH} 或半分子 \ce{FADH2})携带高能电子进入电子传递链,以及一分子 \ce{ATP} 或 \ce{GTP} 直接可用的能量。
- 两分子 \ce{CO2}:代谢废弃物。两个碳原子以 \ce{CO2} 的形式离开循环,二氧化碳分子数与乙酰辅酶 \ce{A} 带入的碳原子数相同,使整个循环的碳原子数守恒。但值得注意的是,这两个碳原子并不是随着乙酰辅酶 \ce{A} 进入循环的那两个碳原子,而是来源于草酰乙酸
调节机制与双向代谢枢纽:
- 精密调节:循环的速度受细胞能量状态(\ce{ATP}/\ce{ADP} 比率)调节。当 \ce{ATP} 充足时,异柠檬酸脱氢酶等会被抑制,减缓生产;反之则被激活。
- 双向代谢:\ce{TCA} 循环不仅是分解途径,产生的中间产物如 \alpha-酮戊二酸和草酰乙酸常被抽走,用于合成氨基酸、脂质和核苷酸等(合成代谢)。
- 添补反应:为了防止中间产物被抽走导致循环停滞,细胞通过添补反应(如丙酮酸羧化为草酰乙酸)来及时补充“燃料”。
这些产生的还原辅酶(\ce{NADH} 和 \ce{FADH2})随后将进入氧化磷酸化阶段。
有氧呼吸的最终阶段:氧化磷酸化
氧化磷酸化是有氧呼吸的最终阶段,也是真核细胞产生 \ce{ATP} 的主要途径。
这一精密过程可以拆解为两个阶段:
电子传递链(ETC)由一系列嵌入线粒体内膜的蛋白质复合体组成。
- 能量来源:来自糖酵解和三羧酸循环的 \ce{NADH} 和 \ce{FADH2} 贡献出高能电子。
- 质子泵送:当电子在复合体间逐级传递时,释放的能量将质子(\ce{H+})从线粒体基质泵入膜间隙。
- 最终受体:电子最终传递给氧气(\ce{O2}),并结合质子生成水(\ce{H2O})。
化学渗透:质子在膜两侧形成了浓度梯度和电位差,这被称为质子动力势。
- \ce{ATP} 合酶:质子顺着电化学梯度通过 \ce{ATP} 合酶回流到基质中。
- 能量转化:\ce{ATP} 合酶如同“分子涡轮”,利用质子流的能量将 \ce{ADP} 和 \ce{Pi} 合成为 \ce{ATP}。
能量统计与调节:
- 高效产出:一分子葡萄糖通过完整氧化分解,最终可产生约 38 个 \ce{ATP} 分子(在理想情况下,实际上大约在 30\sim32 个左右较为合理)。
- 抑制与调节:氰化物通过抑制细胞色素 c 氧化酶阻断电子传递;而解偶联蛋白(如在棕色脂肪中)能使质子不经 \ce{ATP} 合酶流回,从而将能量直接转化为热能。
一些广为人知的药物和毒素可以抑制氧化磷酸化。尽管这些毒素的任何一个都只抑制电子传递链中的一个酶,抑制此过程中的任何步骤都将使其剩余部分停止运转。例如,如果抑制了 \ce{ATP} 合酶,质子就不能回流到线粒体内。结果,质子泵就无法工作,因为梯度差太大以至于无法克服。然后,\ce{NADH} 不再被氧化,而因为 \ce{NAD+} 浓度低于酶可用的浓度,三羧酸循环也随之停止运转。
| 化合物 | 作为 | 作用 |
|---|---|---|
| 氰化物、一氧化碳、叠氮化物、硫化氢 | 毒素 | 抑制细胞色素 c 氧化酶阻断电子传递链 |
| 寡霉素 | 抗生素 | 抑制 \ce{ATP} 合酶 |
| 羰基氰化物间氯苯腙(CCCP)、2,4-二硝基苯酚(DNP) | 毒素 | 携带质子跨膜的离子载体,从而破坏质子梯度。这种离子载体将质子泵与 \ce{ATP} 合酶解耦,因为它能承载质子跨过线粒体内膜。 |
| 鱼藤酮、丙二酸盐、草酰乙酸、抗霉素 A | 农药或毒素 |
并非所有的氧化磷酸化抑制剂都是毒物。在棕色脂肪组织里,称为解偶联蛋白的调节性质子通道可将呼吸作用与 \ce{ATP} 合成解耦。它能使呼吸作用快速产生热量,对冬眠动物和幼年哺乳动物来说,这对维持体温尤为重要,虽然这些蛋白质可能在细胞的应激反应中有更一般化的功能。